Рибонуклеиновые кислоты отличаются от ДНК по химическому составу, строению, функциям, выполняемым в клетке. Молекула РНК, в отличие от ДНК, содержит лишь одну полинуклеидную цепочку. В состав рибонуклеотида (мономер) входят сахар (рибоза), остаток фосфорной кислоты и одно из 4х азотистых оснований (А,Г,Ц,У). в клетке обнаружено несколько типов РНК, три из которых принимают участие в синтезе полипептидов.
Т=У, катализирует химические реакции, первичная структура – нуклеозиды связываются 3' с 5' фосфодиэфирными связями. Первичная структура – одноцепочечная, спирализованная. Третичная – еще раз спирализуется + водородные связи.
и(м)РНК – могут содержать от 300 до 3тыс рибонуклеотидов и имеют линейную структуру. Являются посредниками между ДНК и полипептидом. В процессе синтеза молекулы иРНК с молекулы ДНК переписывается информация о структуре пептида. Далее иРНК переносит информацию из ядра в цитоплазму к рибосомам, где и происходит синтез полипептида. иРНК составляет примерно 0,5-1% массы всех РНК клетки.
рРНК – содержат 3-6тыс рибонуклеотидов, эти молекулы имеют петельную структур. Образуя комплекс с белками, рРНК формируют субъединицы рибосом. Примерно 90% массы всех РНК.
тРНК – переносчик, имеет характерную пространственную структуру, напоминающую кленовый лист или трилистник. Была расшифрована в 1965 году в лаборатории Р.Холлии. В составе 50-100 нуклеотидов. 10-15% массы всей РНК. Функция переноса аминокислот из цитоплазмы к рибосомам, где осущ синтез белка. Первичная структура – 74-95 нуклеотидных остатков. Начало – 3' конец (цитозин+аденозин, ССА-конец). Именно 3' конец аденозин связывается с аминокислотным остатком при образовании аминоацетил-тРНК. ССА-конец присоединяется ко многим тРНК с помощью специального фермента. В других случаях он считывается с кодирующего данную тРНК гена. Нуклеотидный триплет, комплементарный кодону для аминокислоты (антикодон), находится приблизительно в середине цепи тРНК. В отдельных положениях последовательности практически у всех видов тРНК встречаются одни и те же (консервативные) нуклеидые остатки. В некоторых положениях могут находиться или только пуриновые, или только пиримидиновые основания (полуконсервативные). Для всех РНК характерно наличие около 25% разнообразных модификационных нуклеозидов (минорные). Они образуются в различных местах молекул, во многих случаях четко определенных, в результате модификации обычных нуклеозидных остатков с помощью специальных ферментов. Всего более 60 модификаций тРНК. Вторичная структура – 5' конец комплементарен участку, близкому к 3'-концу цепи, при их антипараллельном расположении. Они формируют так называемый акцепторный стебель. В результате образуется структура, характеризующаяся наличием четырех стеблей и трех петель, которая получила название «клеверного листа». Стебель с петлей формируют ветвь, внизу расположена антикодоновая ветвь, содержащая антикодоновый триплет в составе своей петли. Слева и справа от нее расположены D-, Т- ветви, соответственно названные так из-за присутствия в их петлях необычных консервативных нуклеозидов дигидроуридина и тимидина. Нуклеотидные последовательности всех изученных тРНК могут быть сложены в аналогичные структуры. В дополнение к трем петлям клеверного листа в структуре тРНК выделяют дополнительную V-петлю. Ее размеры 4-21 нуклеотида или до 24. Двутяжевые стебли, имеющие постоянное число спаренных нуклеотидов, представляют собой двойную спираль. На виток этой спирали приходится 1 пар нуклеотидных остатков, она близка по параметрам к А-форме ДНК. Неспаренные участки молекулы тРНК (петли и ССА-конец) также имеют вторичную структуру: несколько расположенных друг за другом нуклеотидных остатков образуют однотяжевую спираль за счет межплоскостных взаимодействий их оснований (стэкинг основания). 
Третичная структура – 1974 год, дрожжевая фенилаланиновая тРНК с помощью рентгеноструктурного анализа ее кристаллов. С тех пор удалось закристаллизовать и расшифровать пространственную структуру еще почти десятка тРНК из дрожжей и кишечной палочки. Общие принципы складывания цепей различных тРНК в компактную третичную структуру оказались универсальными. За счет взаимодействия элементов вторичной структуры формируется третичная структура, которая получила название L-формы из-за сходства с латинской буквой L. За счет стэкинга оснований акцепторный стебель и Т-стебель клеверного листа образуют одну непрерывную двойную спираль(одну из палочек доменов буквы L), а два других стебля – антикодоновый и D-другую непрерывную двойную спираль (второй домен L). При этом D- и Т-петли оказываются сближенными и скрепляются между собой путем образования дополнительных, часто необычных пар оснований. В образовании этих пар. Как правило, принимают участие консервативные или полуконсервативные остатки. В свете этого становится ясно, зачем они присутствуют в D- и Т-петлях. Аналогичные третичные взаимодействия скрепляют и некоторые другие участки L-структуры. ССА-конец тРНК и ее антикодоновый триплет находятся на максимальном удалении один от другого, причем основания антикодона обращены внутрь угла L-образной молекулы. Индивидуальные различия в первичной структуре отдельных видов тРНК проявляют себя на уровне как вторичной, так и третичной структур. Известные структуры тРНК, специфичных для разных аминокислот, различаются величиной угла между доменами L-структуры и некоторыми другими деталями.
Вирусы. Двуниточные РНК. Это необычные для клетки типы нуклеиновых кислот, впервые обнаружены у реновирусов, широко распространены среди вирусов животных, растений, бактерий. Вирусы, содержащие подобный геном, назыв диплорнавирусы – у них одноцепочечная РНК, но каждая ее частица имеет 2 копии. +характерной особенностью является обратная транскрипция от РНК к ДНК. После проникновения вируса в клетке вирусный геном начинает обратно транскрибировать, образовывая дуплекс РНК-ДНК и в итоге формирует 2хцепочечную ДНК с помощью фермента – обратной транскриптазы, который содержится в варионах и активизируется после попадания в клетку хозяина, фермент осуществляет синтез, используя ДНК как матрицу.
Репликация ДНК
Репликация (возобновление) – процесс синтеза дочерней молекулы ДНК на матрице родительской молекулы ДНК. Наблюдения показали, что репликация начинается не с конца линейной молекулы, а с образования репликативного глаза, где две цепи родительской ДНК отделяются друг от друга, и в местах расхождения полинуклеотидных цепочек начинается синтез дочерних цепей. Это приводит к появлению в молекуле ДНК петли с виде участка, расплетенного на две отдельные нити.

Участок нуклеиновой кислоты, с которого начинается репликация называется точкой инициации репликации или ориджином. Этот участок содержит последовательность, состоящую из 300 нуклеотидов. Область расхождения двойной спирали ДНК на одинарных полинуклеотидные цепочки в зонах репликации называют репликативной вилкой. Постепенное разъединение цепей молекулы ДНК происходит при разрыве водородных связей между пуринами и пиримидинами. Репликация может происходить в одном или двух направлениях. При однонаправленной репликации вдоль ДНК движется одна реплекационная вилка. При двунаправленной от точки начала в противоположные стороны расходятся две вилки. На определенном расстоянии от точки инициации молекула ДНК содержит участок, в котором репликация останавливается (точки терминации репликации). Участок от точки инициации до точки ее окончания – репликон – единица репликации. Для осуществления репликации цепи материнской ДНК должны быть отделены друг от друга, чтобы стать матрицами, на которых будут синтезироваться комплементарные цепи дочерних молекул. Такое разделение комплементарных полинуклеотидных цепочек происходит с помощью фермента геликазы. Геликаза, разделяя спирально закрученные цепи материнской ДНК, вызывает появление супервитков (те сильную спирализацию молекулы ДНК) перед репликационной вилкой. Супервитки возникают потому, что при расхождении каждых 10 пар нуклеотидов, образующих один виток спирали, материнская ДНК должна совершить один полный оборот вокруг своей оси. Следовательно, для продвижения репликационной вилки вперед вся молекула перед ней должна была бы быстро вращаться. Такое вращение потребовало бы большой затраты энергии. В действительности же вращения не происходит благодаря действию особого фермента – ДНК-топоизомеразы. Топоизомераза разрывает одну из цепей ДНК и совершает оборот вокруг второй (целой) цепи как вокруг оси вращения. 
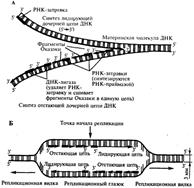
Это устраняет супервитки и ослабляет накопившиеся напряжение в двойной спирали ДНК. После снятия избыточного напряжения фермент восстанавливает разорванную цепь. К высвобождающимся водородным связям нуклеотидных последовательностей разделенных родительских цепей присоединяются свободные нуклеотиды из нуклеоплазмы, где они присутствуют в виде дезоксирибонуклеозидгрифосфатов: дАТФ, дГТФ, дЦТФ, дТТФ. Комплементарный нуклеозидтрифосфат образует водородные связи с определенным основанием материнской цепи ДНК. Затем при участии фермента ДНК-полимеразы он связывается фосфодиэфирной связью с предшествующим нуклеотидом вновь синтезируемой цепи, отдавая при этом неорганический пирофосфат. Поскольку ДНК-полимераза присоединяет очередной нуклеотид к ОН-группе в 3'-положении предшествующего нуклеотида, цепь постепенно удлиняется на ее 3'-конце. Особенностью ДНК-полимеразы является ее неспособность начать синтез новой полинуклеотидной цепи путем простого связывания двух нуклеозидтрифосфатов: необходим 3'-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинук-леотиднуй цепь называют затравкой или праймером. Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы (рис. 3.11). Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3'-ОН-конец. Способность ДНК-полимеразы осуществлять сборку полинуклеотида в направлении от 5'- к 3' -концу при антипараллельном соединении двух цепей ДНК означает, что процесс репликации должен протекать на них по-разному. Действительно, если на одной из матриц (3' → 5') сборка новой цепи происходит непрерывно от 5'- к 3'-концу и она постепенно удлиняется на 3'-конце, то другая цепь, синтезируемая на матрице (5' → 3'), должна была бы расти от 3'- к 5'-концу. Это противоречит направлению действия фермента ДНК-полимеразы. В настоящее время установлено, что синтез второй цепи ДНК осуществляется короткими фрагментами (фрагменты Оказаки) также в направлении от 5'- к 3'-концу (по типу шитья «назад иголкой»). У прокариот фрагменты Оказаки содержат от 1000 до 2000 нуклеотидов, у эукариот они значительно короче (от 100 до 200 нуклеотидов). Синтезу каждого такого фрагмента предшествует образование РНК-затравки длиной около 10 нуклеотидов. Вновь образованный фрагмент с помощью фермента ДНК-лигазы соединяется с предшествующим фрагментом после удаления его РНК-затравки (рис. 3.12, А). В связи с указанными особенностями репликационная вилка является асимметричной. Из двух синтезируемых дочерних цепей одна строится непрерывно, ее синтез идет быстрее и эту цепь называют лидирующей. Синтез другой цепи идет медленнее, так как она собирается из отдельных фрагментов, требующих образования, а затем удаления РНК-затравки. Поэтому такую цепь называют запаздывающей (отстающей). Хотя отдельные фрагменты образуются в направлении 5' → 3', в целом эта цепь растет в направлении 3' → 5' (рис. 3.12, А). В виду того, что от локуса ori как правило начинаются две репликационные вилки, идущие в противоположных направлениях, синтез лидирующих цепей в них идет на разных цепях материнской ДНК (рис 3.12, Б). Конечным результатом процесса репликации является образование двух молекул ДНК, нуклеотидная последовательность которых идентична таковой в материнской двойной спирали ДНК. Рассмотренная последовательность событий, происходящих в ходе репликативного синтеза, предполагает участие целой системы ферментов: геликазы, топоизомеразы, дестабилизирующих белков, ДНК-полимеразы и других, совместно действующих в области репликационной вилки (рис 3.13). Репликация ДНК у про- и эукариот в основных чертах протекает сходно, однако, скорость синтеза у эукариот (около 100 нуклеотидов/с) на порядок ниже, чем у прокариот (1000 нуклеотидов/с). Причиной этого может быть образование ДНК эукариот достаточно прочных соединений с белками (см. гл 3.5.2.), что затрудняет ее деспирали-зацию, необходимую для осуществления репликативного синтеза. Фрагмент ДНК от точки начала репликации до точки ее окончания образует единицу репликации — репликон. Однажды начавшись в точке начала (локус on), репликация продолжается до тех пор, пока весь репликон не будет дуплицирован. Кольцевые молекулы ДНК прокариотических клеток имеют один локус on и представляют собой целиком отдельные репликоны. Эукариотические хромосомы содержат большое число репликонов. В связи с этим удвоение молекулы ДНК, расположенной вдоль эукариотической хромосомы, начинается в нескольких точках. В разных репликонах удвоение может идти в разное время или одновременно.






