Экспрессия гена представляет собой процесс реализации закодированной в гене информации в виде признака - фенотипа. На клеточном уровне она заключается в последовательности этапов, в результате которых записанная в ДНК информация реализуется путем формирования признаков. На молекулярном уровне это процесс реализации генетической информации в виде полипептида, рРНК или тРНК.
Экспрессия генов является сложным процессом и включает следующие этапы:
→ активация и транскрипция гена - переписывание генетической информации, с ДНК и синтез первичных транскриптов (гетерогенная ядерная РНК);
→ процессинг - созревание мРНК: кэпирование, полиаденилирование, сплайсинг;
→ перенос мРНК в цитоплазму;
→ трансляция — процесс перевода генетической информации из нуклеотидной последовательности в последовательность аминокислот полипептидной цепи;
→ конформация - преобразование полипептидной цепи в белок.
На клеточном уровне экспрессия гена является результатом интеграции вновь синтезированного белка в определенную клеточную структуру, цепь метаболизма или в систему клеточной сигнализации. Фенотип клетки - морфология и функции - определяются ее геномом и реализуется при участии целого набора синтезированных клеткой белков - протеосома. Специфичность белков является основой дифференцировки и специализации клеток.
На уровне организма экспрессия гена проявляется в виде различных признаков и свойств благодаря взаимодействию молекулярных и надмолекулярных компонентов в процессе морфогенеза и развития организма человека.
Следовательно, в рамках современной концепции гена выделяют три уровня экспрессии генов:
- молекулярный - заключается в синтезе полипептида, который является первичным продуктом экспрессии генов;
- клеточный - конформация и образование функциональных белков, выполняющих различные функции в клетке;
- организменный - фенотипическое проявление гена в виде признака.
Например, мутация гена β-глобина является причиной серповидно-клеточной анемии. На молекулярном уровне это проявляется в виде синтеза дефектной полипептидной цепи, в которой глутаминовая кислота заменена на валин. Как следствие, вместо нормального гемоглобина НЬА образуется мутантный гемоглобин HbS. На клеточном уровне наблюдается образование аномальных эритроцитов серповидно-клеточной формы. Это приводит к проявлению патологических признаков анемии на уровне организма.
КЛАССИФИКАЦИЯ ГЕНОВ ЧЕЛОВЕКА
Различают несколько классификаций генов, основанных на разных критериях. Например:
1. по типу конечного продукта:
- гены, кодирующие белки - структурные гены;
- гены, кодирующие рРНК и тРНК;
2. по количеству копий в геноме:
- уникальные - представлены, как правило, одной копией;
- повторяющиеся - организованы тандемно или разбросаны по геному;
3. по количеству клеток, в которых гены активны:
- общие гены,или гены домашнего хозяйства;
- тканеспецифичные гены.
4. по времени фенотипического проявления:
- активные в эмбриональном периоде;
- активные на этапе полового созревания;
- активные во взрослом организме.
5. по степени активности:
- нормоморфные;
- гипоморфные;
- гиперморфные;
- аморфные.
Об активности генов можно судить по количеству транскрибированной РНК и количеству синтезированного белка.
6. по функции конечного продукта:
- ферменты - 31,2%;
- модуляторы функции белков -13, 6 %
- рецепторы;
- факторы транскрипции;
- белки внутриклеточного и внеклеточного матриксов; 50%
- транспортные белки;
- сигнальные молекулы;
- гормоны;
- иммуноглобулины.
7. по зависимости экспрессии генов от негенетических факторов:
- стабильные;
- пластичные.
ЛОКАЛИЗАЦИЯ ГЕНОВ
Согласно хромосомной теории наследственности Т.Моргана (1911): (1) гены расположеныв хромосомах в линейном порядке, занимая определенное место - локус; (2) гены одной хромосомы образуют одну группу сцепления и наследуются вместе; (3) количество групп сцепления равно гаплоидному числу хромосом; (4) между гомологичными хромосомами может происходить обмен участками - кроссинговер (5) частота кроссинговера прямо пропорциональна расстоянию между генами и обратно пропорциональна силе сцепления; (6) расстояние между генами измеряется в сантиморганидах - 1сМ = 1% кроссинговера.

Гены, расположенные в идентичных локусах гомологичных хромосом выполняют одинаковые функции и называются аллельными, в то время как гены, расположенные в разных локусах гомологичных или негомологичных хромосом, называются неаллелъными
.

Распределение генов вдоль хромосом неодинаково: хромосомы отличаются по количеству и плотности расположения генов. Некоторые гены представлены в одном экземпляре, а другие - многими копиями и образуют семейства повторяющихся и неповторяющихся генов.
Гены одной хромосомы, расположенные близко друг от друга, образуют гаплотип и имеют часто общие регуляторные элементы. В широком смысле под гаплотипом понимают совокупность генов одной молекулы ДНК.
Гены, локализованные в аутосомах, определяют аутосомные признаки и наследуются независимо от пола, а гены, расположенные в гоносомах, определяют сцепленные с полом признаки, и их наследование зависит от пола:
- Х-сцепленные гены и признаки передаются от матери к дочерям и сыновьям, а от, отца - только дочерям;
- Y-сцепленные гены и признаки (голандрические) передаются исключительно от отца к сыну.
ГЕНЕТИЧЕСКИЕ КАРТЫ
В состав генома человека входят две различающиеся по организации и особенностям наследования системы: ядерный геном и митохондриальный геном. В ядре соматических клеток человека содержится около 30 000 пар генов, которые локализованны в 46 молекулах ДНК.
Каждая хромосома содержит в среднем 2000 генов. Гены расположены в линейном порядке и разделены некодирующими последовательностями (сателлитная ДНК, спейсеры). Гены одной хромосомы наследуются совместно. Это явление называется сцеплением генов. Каждая хромосома, таким образом, представляет одну группу сцепления. Митохондриальный геном представлен кольцевыми молекулами ДНК и содержит 37 генов, расположенными очень компактно и наследуемыми по материнской линии.

Явление сцепление характерно только для генов одной и той же хромосомы, в то время как расположенные в разных хромосомах гены наследуются независимо, по законам Менделя. Сцепление бывает полным и неполным. Причина неполного сцепления - кроссинговер, происходящий в мейозе. В ходе кроссинговера происходит взаимный обмен аллельными генами.
Частота кроссинговера неодинакова для различных локусов и может варьировать от 0% до 50%, коррелируя с расстоянием между генами. Чем далее друг от друга расположены гены, тем больше частота кроссинговера между ними и, наоборот. Данное положение позволяет определить расстояние между генами по частоте кроссинговера и лежит в основе составления генетических карт. Эти карты представляют собой графическое изображение хромосом и расположенных на них генов с указанием расстояния между ними.

В настоящее время, благодаря методам дифференциальной окраски и соматической гибридизации, разработаны физические карты хромосом, в которых показано точное расположение генов с указанием расстояния между ними в парах нуклеотидов.
Установление сцепления между генами и определение групп сцепления представляет значительный интерес в медицинской генетике. Примерами сцепленных генов у человека являются: ген фактора Rh и ген эллиптоцитоза; ген АВО и ген пигментной ксеродермы (ХР); гены Duffy и врожденной катаракты; ген MNSs и dentinogenesis imperfecta-1 (DI-1); ген Xg и гены гемофилии А (НЕМА), гемофилии В (НЕМВ), дальтонизмаI (Dalt).

МЕТОДЫ АНАЛИЗА ГЕНОВ
Методы рекомбинантной ДНК создали предпосылку для разработки новых методов молекулярной диагностики с большей разрешающей способностью и, следовательно, более точных и информативных. Об абсолютном преимуществе молекулярного подхода говорит тот факт, что в отличие от других методов диагностики, ограниченных выявлением исключительно фенотипических аспектов, анализ ДНК, направленный непосредственно на изучение генотипа, является единственным методом, изучающим первичные нарушения (мутации), то есть именно первопричину возникновения болезней.
Технология рекомбинантной ДНК позволяет определить нормальные гены и/или мутантные их варианты, установить носителей мутантного гена, диагностировать наследственную патологию до рождения ребенка или в предсимптоматической стадии, а в ближайшем будущем проводить и генную терапию.
Молекулярное изучение генов может осуществляться многими путями в зависимости от поставленной цели:
1. секвестрование ДНК для определения первичной структуры гена;
2. метод Саузерн-блот для определения позиции гена в геноме;
3. метод Нозерн-блот для определения экспрессии генов (анализ мРНК);
4. метод Вестерн-блот для определения белкового продукта гена;
5. метод ПЦР для специфичной амплификации фрагментов ДНК.
В лабораториях молекулярной биологии используются различные варианты перечисленных методов.
Все перечисленные методы основаны на различных принципах манипулирования нуклеиновыми кислотами:
- специфическая рестрикция ДНК с целью получения интересующего фрагмента;
- идентификация фрагментов ДНК или РНК е помощью специальных зондов, комплементарных искомому участку;
- идентификация нормальных и патологических генов путем ПЦР - специфическая реакция отбора праймеров, комплементарных гену/последовательности;
- визуализация необходимого фрагмента по результатам электрофореза и специфического маркирования ДНК/РНК с использованием компьютерных программ чтения и анализа результатов;
- комплексная интерпретация результатов в зависимости от использованного метода.
| Для разделения полученных фрагментов ДНК применяется электрофорез макромолекул в агарозном или полиакриламидном геле. Будучи отрицательно заряжены, фрагменты молекул нуклеиновых кислот в электрическом поле перемещаются с разной скоростью в зависимости от молекулярной массы. Более короткие фрагменты перемещаются быстрее, в то время, как более длинные - медленнее. Для определения размеров фрагментов в геле, одновременно с интересующими фрагментами помещаются на соседние дорожки и фрагменты - маркеры длины. Молекулы нуклеиновых кислот могут быть обнаружены в геле при окраске флуоресцентным красителем или по радиоактивной метке. В случае радиоактивного мечения, фрагменты определяются с помощью авторадиографии, которая заключается в накладывании на гель светочувствительной пленки.
|

Секвенирование ДНК
Секвенирование заключается в определении последовательности нуклеотидов (азотистых оснований) в исследуемом фрагменте ДНК. Анализ последовательностей нуклеотидов ДНК может быть осуществлен двумя путями: 1) химическим (метод Максима - Гильберта), при котором используются химические реакции последовательного расщепления фрагмента ДНК на отдельные нуклеотиды, однако этот метод очень трудоемкий, сложный и в последнее время не используется; 2) ферментативным (метод Сэнгера), при котором фрагменты ДНК синтезируются in vitro таким образом, что реакция завершается в положении, соответствующем определенному основанию. Для определения последовательности нуклеотидов любым из этих методов, ДНК подвергается серии из 4-х отдельных реакций, каждая из которых является специфической для одного из 4-х видов нуклеотидов. При одновременном электрофорезе продукты реакции на 4-х соседних дорожках будут мигрировать по ним с разной скоростью. Размер синтезированных фрагментов можно определить, а, следовательно, может быть определен и порядок нуклеотидов в
ДНК.
Метод Сэнгера (дидеокси) использует ферментативный синтез одной цепи, комплементарной клонированной матрице. В ходе данного процесса синтез ДНК останавливается добавлением одного из дидезоксинуклеозидтрифосфата, аналога нуклеотидов. Дидезоксинуклеозидтрифосфат содержит в позиции 3' не группу ОН, а Н-группу, которая препятствует полимеризации нуклеотидов. Используя четыре разных аналога дидезокси в процессе синтеза новой цепи ДНК, можно, таким образом, определить каждый нуклеотид матричной цепи. Электрофорез полученных фрагментов позволяет определить порядок нуклеотидов ДНК.
В последние годы применяются методы автоматического секвенирования.
С целью диагностики носителей мутантного гена результат секвенирования сравнивают с первичной структурой гена. К сожалению, не для всех генов человека к настоящему моменту известны последовательности нуклеотидов, поэтому применяются и непрямые методы диагностики: сцепление с близлежащими ДНК-маркерами (определение гйпервариабельных мини- и микросателлитных повторов), определение характерных для данного гена полиморфных сайтов рестрикции, гибридизация со специфическими зондами и др.

Метод Саузерн-блот
Метод основан на специфическом анализе определенных фрагментов геномной ДНК/гена, полученных путем рестрикции ДНК одной или несколькими рестриктазами. Ферменты рестрикции действуют не случайно, а разрезают ДНК в строго определенных местах, поэтому в результате действия одной рестриктазой получают двухцепочечные фрагменты ДНК разной длины (количество фрагментов и их длина специфичны для данной рестриктазы). Учитывая полиморфизм ДНК/генов, обусловленный точечными мутациями, длины фрагментов специфичны и неодинаковы у разных индивидов (полиморфизм длины рестрикционных фрагментов - ПДРФ).
Для анализа ПДРФ ряда генов необходимо знать специфическую локализацию сайтов рестрикции. Эта информация может быть полезна для сравнения нормальных и мутантных генов для раннего выявления носителей патологических генов. На данном принципе основан метод Саузерн-блот, который позволяет идентифицировать интересующий фрагмент в смеси из тысячи разных фрагментов, полученных в результате рестрикции геномной ДНК. Идентификация необходимого фрагмента осуществляется на основе гибридизации ДНК-мишени с комплементарным меченым зондом.
Метод Саузерн-блота состоит из последовательности следующих этапов:
(1) выделение высокомолекулярной геномной ДНК из клетки;
(2) ферментативное расщепление ДНК разными рестриктазами с образованием фрагментов разной длины;
(3) разделение рестрикционных фрагментов в агарозном геле;
(4) денатурация фрагментов ДНК щелочным раствором;
(5) нейтрализация геля буферным раствором;
(6) капиллярный перенос фрагментов на нейлоновый или нитроцеллюлезный фильтр;
(7) гибридизация с одноцепочечными радиоактивными зондами;
(8) авторадиография для выявления гибридов: исследуемая ДНК / зонд.
Перенос фрагментов на нитроцеллюлезный фильтр - блоттинг - предложил Эдвард Саузерн.
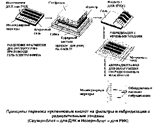
В результате анализа полученного материала можно определить наличие или отсутствие некоторых сайтов рестрикции, характерных для изучаемого гена, которые ассоциируются с определенными мутациями. Различные варианты расположения сайтов рестрикции в ДНК у двух разных людей называют Полиморфизмом Длины Рестрикционных Фрагментов (ПДРФ). Этот полиморфизм используется как генетический маркер в изучений генотипа.
Практическое применение метода Саузерн-блот:
- выявление точечных мутаций, затрагивающих сайты рестрикции (появляются новые или исчезают старые сайты рестрикции) по изменению количества и длины фрагментов рестрикции;
- выявление мутаций типа делении, дупликации, инсерции более длинных фрагментов (50-100 п.н.) по изменению длины фрагментов рестрикции.
Это позволяет проводить пренатальную и пресимптоматическую диагностику патологических мутаций и выявление гетерозиготных носителей мутантных генов.
Метод Саузерн-блот имеет, однако, следующие недостатки: (1) не позволяет выявить точечные мутации, не затрагивающие сайтов рестрикции (в пределах фрагмента рестрикции); (2) является достаточно трудоемким, сложным и дорогим методом.
Метод Нозерн-блот
Метод состоит в переносе разделенных молекул РНК на нейлоновые или нитроцеллюлезные фильтры с последующей гибридизацией с мечеными зондами. Метод схож с методом Саузерн-блот, за исключением того, что выделенные и очищенные мРНК не подвергают рестрикции, а электрофорез не происходит в условиях денатурации.
Метод Нозерн-блот позволяет идентифицировать транскрипты анализируемых генов, количества мРНК, их размеры.
Метод Бестерн-блот
Этот метод заключается в определении специфического белка из смеси клеточных белков. Для этого белки разделяют электрофорезом в условиях денатурации, в присутствии додецилсульфатa Na. Белки, разделённые по молекулярной массе, переносят на плотный фильтр и обрабатывают специфическими мечеными антителами. Этот метод позволяет определить наличие или отсутствие белка, его размер и скорость экспрессии гена по количеству синтезированного белка.
Техника ПЦР в анализе генов
Метод ПЦР используется для выборочного размножения определенной последовательности гена. В результате получают гомогенные популяции фрагментов, которые используются в исследованиях по молекулярной генетике или диагностике. Для осуществления амплификации определенной последовательности необходимо знать первичную структуру нормального или мутантного гена и синтезировать специфические праймеры, комплементарные концам интересующих фрагментов. Праймеры представляют собой искусственно синтезированные одноцепочечные олигонуклеотиды (20-30 нуклеотидов). ПЦР основана на гибридизации исследуемой последовательности ДНК - праймер и полуконсервативной репликации ДНК.
Преимущества метода ПЦР следующие: требуются минимальные количества ДНК; высокая скорость амплификации (за несколько часов получают более миллиона копий одинаковых фрагментов); благодаря специфичности праймера амплифицируетея только нужный фрагмент, который затем используется как зонд в других методах.







