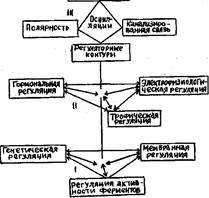
ОСОБЕННОСТИ МЕЖКЛЕТОЧНОЙ РЕГУЛЯЦИИ
Трофическая рег-я – взаимодействие с помощью питательных веществ – наиболее простой способ связи между клетками, тканями и органами; носит колличественный, а не качественный характер.
Гормональная рег-я – важнейший фактор регуляции и управления у растений. Фито- гормоны – ауксин (индолил-3-уксусная кислота)- растяжение кл., цитокинины (зеатин, изопентениладенин)- деление кл., гиббереллины- удлинение побега, абсцизовая кислота, этилен(ингибиторы роста,стресс-гормоны) – сравнительно низкомолекулярные органические вещества с высокой физиологической активностью, присутствующие в тканях в очень низких концентрациях, с помощью которых клетки, ткани и органы взаимодействуют между собой.
Электрофизиологическая рег-я - включает в себя возникновение градиен тов биоэлектропотенциалов (БЭП) между разными частями растения и генерацию распространяющихся потенциалов (потенциала действия и вариабельного потенциала). Градиенты БЭП возникают благодаря различию величин мембранного потенциала (МП) в клетках разных тканей, зон и органов растительного организма. Эти градиенты не остаются постоянными, а совершают медленные периодические колебания, обусловленные изменениями условий внутренней и внешней среды.
Межклеточные системы регуляции на клеточном уровне действуют только через внутриклеточные системы регуляции, то есть через метаболическую, генетическую и мембранную системы. Таким образом осуществляется принцип иерархии систем регуляции в целом растительном организме.
Опишите механизм фотофосфолирования
Это присоединение фосфатов с выделением АТФ. Энергия поглащения хлорофиллом при переносе электрона от одного переносчика к другому обеспечивает преобразование в энергию химических связей. Таким образом фотофосфолирование это синтез АТф протекающий с использованием видимой части солнечной радиации. Впервые предложил эту теорию Митчел.
Редукция нитрата
В органические соединения включается только аммонийный азот, поэтому ионы нитрата, поглощенные растением, восстанавливаются в клетках до аммиака. Редукция нитрата в растениях осуществляется в два этапа.
Сначала происходит восстановление нитрата до нитрита, сопряженное с переносом 2 электронов и
катализируемое ферментом нитратредуктазой:
2 e-
NO-3 + НAД(Ф)Н + Н+ NO-2 + НАД(Ф)+ + Н2О
Никотинамидадениндинуклеотидфосфат восстановленный (НАДФН). никотинамидадениндинуклеотиду восстановленному (НАДН), который образуется в ходе реакий
Нитриты, образующиеся на первом этапе редукции нитратов, быстро восстанавливаются до аммиака ферментом нитритредуктазой. Она в качестве донора электронов использует восстановленный ферредоксин:
6 е-
NO-2 + 6 Фдвосст. + 8 Н+ NH+4 + 6 Фдокисл. + 2 Н2О
Обе эти реакции происходят в листьях и корнях. В зеленых частях растения нитритредуктаза локализована в хлоропластах. Восстановитель ферредоксин получает электроны прямо из фотосинтетической электронтранспортной цепи. В корнях нитрит восстанавливается в пропластидах. Так как в корнях ферредоксин отсутствует, то источником электронов служит НАДФН, образующийся в пентозофосфатном пути дыхания.
Пути ассимиляции аммиака
Аммиак, поступивший в растение из почвы, образовавшийся при восстановлении нитратов или в процессе фиксации молекулярного азота, усваивается растениями с образованием аминокислот и амидов. Фермент глутаматдегидрогеназа катализирует восстановительное аминирование -кетоглутаровой кислоты с образованием глютаминовой кислоты. На первом этапе реакции субстраты соединяются с образованием аминокислоты, которая затем восстанавливается в глютаминовую кислоту при участии НАД(Ф)Н.
Глютаматдегидрогеназа обнаружена в листьях и корнях у всех высших растений, но в корнях ее активность выше. Фермент локализован преимущественно в митохондриях, хотя имеется в цитоплазме и в хлоропластах.
Глютаминсинтетаза катализирует реакцию, в которой глютаминовая кислота функционирует как акцептор NH3 для образования амида глютамина. Ионы марганца, кобальта, кальция и магния являются кофакторами глютаминсинтетазы. Фермент обнаружен во всех органах растений и локализован в цитоплазме.
Помимо -кетоглутаровой кислоты, играющей основную роль в первичном связывании аммиака, роль акцепторов аммиака в растениях могут выполнять и другие органические кислоты, которые с помощью соответствующих ферментов взаимодействуют с NH3, образуя так называемые первичные аминокислоты. Они же служат акцептором аминогрупп в различных реакциях переаминирования. К числу этих органических кислот относятся щавелевоуксусная, пировиноградная, гидроксипировиноградная, глиоксиловая и другие, в процессе восстановительного аминирования которых получаются соответственно аспарагиновая кислота, аланин, серин, глицин.
Принято считать, что образование аспарагина преобладает в том случае, когда происходит распад белков в семенах. В клетках корня и листьев растущего растения идет, главным образом, образование глютамина. Таким образом, образование аспарагина - это путь обезвреживания аммиака, появляющегося при распаде белка - так называемая регрессивная ветвь азотного обмена, тогда как синтез глютамина - это путь обезвреживания аммиака при синтезе белка - прогрессивная ветвь азотного обмена.
ОСОБЕННОСТИ МЕЖКЛЕТОЧНОЙ РЕГУЛЯЦИИ
Трофическая рег-я – взаимодействие с помощью питательных веществ – наиболее простой способ связи между клетками, тканями и органами; носит колличественный, а не качественный характер.
Гормональная рег-я – важнейший фактор регуляции и управления у растений. Фито- гормоны – ауксин (индолил-3-уксусная кислота)- растяжение кл., цитокинины (зеатин, изопентениладенин)- деление кл., гиббереллины- удлинение побега, абсцизовая кислота, этилен(ингибиторы роста,стресс-гормоны) – сравнительно низкомолекулярные органические вещества с высокой физиологической активностью, присутствующие в тканях в очень низких концентрациях, с помощью которых клетки, ткани и органы взаимодействуют между собой.
Электрофизиологическая рег-я - включает в себя возникновение градиен тов биоэлектропотенциалов (БЭП) между разными частями растения и генерацию распространяющихся потенциалов (потенциала действия и вариабельного потенциала). Градиенты БЭП возникают благодаря различию величин мембранного потенциала (МП) в клетках разных тканей, зон и органов растительного организма. Эти градиенты не остаются постоянными, а совершают медленные периодические колебания, обусловленные изменениями условий внутренней и внешней среды.
Межклеточные системы регуляции на клеточном уровне действуют только через внутриклеточные системы регуляции, то есть через метаболическую, генетическую и мембранную системы. Таким образом осуществляется принцип иерархии систем регуляции в целом растительном организме.








