Дыхательная электронтранспортная цепь — система структурно и функционально связанных трансмембран-ных белков и переносчиков электронов. ЭТЦ позволяет запасти энергию.Компоненты дыхательной цепи: включает три белковых комплекса (комплексы I, III и IV), встроенных во внутреннюю митохондриальную мембрану, и две подвижные молекулы-пе-реносчики — убихинон (кофермент Q) и цитохром. Ком-плексы дыхательной цепи построены из множества полипептидов и содержат ряд различных окислительно-восстановительных коферментов, связанных с белками. Компоненты дыхательной цепи погружены в двойной липидный слой.
41) ХЕМИОСМОТИЧЕСКАЯ ТЕОРИЯ учение о механизме преобразования энергии в биол. мембранах при синтезе аденозинтрифосфорной к-ты (АТФ). Разработана П. Митчеллом в 1961—66. Согласно исходным представлениям Митчелла, запасание энергии в АТФ происходит вследствие предварит, накопления зарядов на стенках мембраны, создания мембранного потенциала и разности концентраций протонов. Разность электрохимич. потенциалов ионов водорода на сопрягающих мембранах (внутр. мембраны митохондрий, тилакоиды хлоропластов, мембраны бактерий) возникает за счёт энергии, выделяемой при деятельности цепи окислит.-восстановит, ферментов, или за счёт поглощённых квантов света. Трансмембранные электрохимич. потенциалы ионов могут служить источником энергии не только для синтеза АТФ, на и для транспорта веществ, движения бактериальных клеток и др. энергозависимых процессов.
42) Каким образом работают механизмы регуляции дыхания у растений.
Важнейшим этапом питания органическими веществами на клеточном уровне является процесс дыхания.
Клеточное дыхание — это окислительный, с участием кислорода распад органических питательных веществ, сопровождающийся образованием химически активных метаболитов и освобождением энергии, которые используются клетками для процессов жизнедеятельности. Дыхание, подобно фотосинтезу, сложный окислительно_восстановительный процесc, идущий через ряд этапов. На его промежуточных стадиях образуются органические соединения, которые затем используются в различных метаболических реакциях. К промежуточным соединениям относят органические кислоты и пентозы образующиеся при разных путях дыхательного распада. Таким образом, процесс дыхания -- источник многих метаболитов. Окисление дыхательных субстратов в ходе дыхания осуществляется с участием ферментов. Ферменты как белковые катализаторы, помимо свойств, присущих неорганическим катализаторам, обладают рядом особенностей: высокой актианостью, высокой специфичностью по отношению к субстратам и высокой лабильностью. Их пространственная организации зависящая от нее активность изменяются под действием внешних и внутренних факторов. Эти свойства обеспечивают возможность тонкой регуляции обмена веществ на уровне ферментов.
Типы окислительно-восстановительных реакций. Существуют четыре способа окисления, и все они связаны с отнятием электронов:
1)непосредственная отдача электронов, например: 
2) Отнятие водорода: 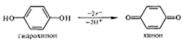
3) присоединение кислорода

4)образование промежуточного гидратированного соединения с последующим отнятием двух электронов и протонов:

43) Перечислите экологические факторы, влияющие на процесс дыхания у растений. Температура. Дыхание у ряда растений осуществляется и при температуре ниже 0°С. Ферментативная реакция с повышением температуры интенсивность дыхания возрастает. Однако это происходит до определенного предела, выше которого начинается инактивация ферментов и интенсивность дыхания снижается. При этом надо учитывать длительность выдерживания растения при данной температуре. При кратковременной экспозиции интенсивность дыхания возрастает при повышении температуры до 35°С и даже 40°С. При длительном выдерживании в такой температуре интенсивность дыхания уменьшается. Снабжениекислородом. Кислород необходим для протекания дыхания, поскольку является конечным акцептором электронов, движущихся по дыхательной цепи. Увеличение содержания кислорода до 5—8% сопровождается повышением интенсивности дыхания. Дальнейшее возрастание концентрации 02 обычно уже не сказывается на интенсивности дыхания. Однако из этого общего положения имеются исключения. Снабжение растительных тканей и клеток кислородом зависит не только от его содержания во внешней среде, но и от скорости его поступления. Между тем часто проникновение кислорода к тем или иным тканям затруднено. Содержание углекислого газа. С02 является конечным продуктом как брожения, так и аэробного дыхания. При довольно высоких концентрациях С02, значительно превышающих те, которые обычно окружают растительный организм (выше 40%), процесс дыхания тормозится. Торможение вызывается несколькими причинами: 1) высокая концентрация С02 может оказывать общее анестезирующее влияние на растительный организм; 2) С02 тормозит активность ряда дыхательных ферментов; 3) повышение содержания С02вызывает закрытие устьиц, что затрудняет доступ кислорода и косвенно тормозит процесс дыхания. Содержание воды. Небольшой водный дефицит растущих тканей увеличивает интенсивность дыхания. Это связано с тем, что водный дефицит и даже подвядание листьев усиливают процессы распада сложных углеводов (крахмала) на более простые (сахара). Увеличение содержания Сахаров (основного субстрата дыхания) усиливает сам процесс. Вместе с тем при водном дефиците нарушается сопряжение окисления и фосфорилирования. Свет. Под влиянием света, особенно коротковолновых сине-фиолетовых лучей, интенсивность обычного темнового дыхания возрастает. Активация дыхания светом показана на бесхлорофилльных растениях. Возможно также, что свет активирует дыхательные ферменты (оксидазы). Питательные соли. Интенсивность дыхания сильно зависит от снабжения растения элементами минерального питания. Такие элементы, как фосфор, сера, железо, медь, марганец, принимают непосредственное участие в процессе дыхания, входя в промежуточные продукты (фосфор) или являясь составной частью дыхательных ферментов. Поранение. Поранение органов и тканей растения усиливает интенсивность дыхания. Это связано с разрушением клеток, из-за чего повышается соприкосновение дыхательных субстратов и ферментов. Частично поранение может вызывать переход клеток в меристематическую фазу роста. Интенсивность дыхания делящихся клеток всегда выше по сравнению с клетками, закончившими рост.
46. Химизм фиксации атмосферного азота.
Конечным продуктом фиксации азота является аммиак. В процессе восстановления азота до аммиака участвует мультиферментный комплекс — нитрогеназа.
Источником протонов и электронов для восстановления азота служит дыхательная электрон-транспортная цепь. Это указывает на связь усвоения азота атмосферы с процессами дыхания, а также фотосинтеза. Для восстановления N2 до NH3 требуется шесть электронов, согласно уравнению:
N2 + 6е + 2Н+ -> 2NH3
по расчетам для восстановления одной молекулы N2 требуется не менее 12 молекул АТФ. Нитрогеназа — фермент восстанавливает и другие соединения с тройной связью: цианиды, ацетилен, азиды и др. Особенность нитрогеназы заключается и в том, что для работы фермента требуются анаэробные условия. Для нормального протекания процесса азотофиксации необходимы Мо и Fe, поскольку они входят в состав фермента нитрогеназы. Молибден выполняет структурную функцию, поддерживая конформацию нитрогеназы, каталитическую, участвуя в связывании азота и переносе электронов, а также индуцирует синтез нитрогеназы. В надземные органы растения-хозяина азотистые вещества передвигаются главным образом в виде амидов (аспарагина, глутамина).
Редукция нитрата
В органические соединения включается только аммонийный азот, поэтому ионы нитрата, поглощенные растением, восстанавливаются в клетках до аммиака. Редукция нитрата в растениях осуществляется в два этапа.
Сначала происходит восстановление нитрата до нитрита, сопряженное с переносом 2 электронов и
катализируемое ферментом нитратредуктазой:
2 e-
NO-3 + НAД(Ф)Н + Н+ NO-2 + НАД(Ф)+ + Н2О
Никотинамидадениндинуклеотидфосфат восстановленный (НАДФН). никотинамидадениндинуклеотиду восстановленному (НАДН), который образуется в ходе реакий
Нитриты, образующиеся на первом этапе редукции нитратов, быстро восстанавливаются до аммиака ферментом нитритредуктазой. Она в качестве донора электронов использует восстановленный ферредоксин:
6 е-
NO-2 + 6 Фдвосст. + 8 Н+ NH+4 + 6 Фдокисл. + 2 Н2О
Обе эти реакции происходят в листьях и корнях. В зеленых частях растения нитритредуктаза локализована в хлоропластах. Восстановитель ферредоксин получает электроны прямо из фотосинтетической электронтранспортной цепи. В корнях нитрит восстанавливается в пропластидах. Так как в корнях ферредоксин отсутствует, то источником электронов служит НАДФН, образующийся в пентозофосфатном пути дыхания.
Пути ассимиляции аммиака
Аммиак, поступивший в растение из почвы, образовавшийся при восстановлении нитратов или в процессе фиксации молекулярного азота, усваивается растениями с образованием аминокислот и амидов. Фермент глутаматдегидрогеназа катализирует восстановительное аминирование -кетоглутаровой кислоты с образованием глютаминовой кислоты. На первом этапе реакции субстраты соединяются с образованием аминокислоты, которая затем восстанавливается в глютаминовую кислоту при участии НАД(Ф)Н.
Глютаматдегидрогеназа обнаружена в листьях и корнях у всех высших растений, но в корнях ее активность выше. Фермент локализован преимущественно в митохондриях, хотя имеется в цитоплазме и в хлоропластах.
Глютаминсинтетаза катализирует реакцию, в которой глютаминовая кислота функционирует как акцептор NH3 для образования амида глютамина. Ионы марганца, кобальта, кальция и магния являются кофакторами глютаминсинтетазы. Фермент обнаружен во всех органах растений и локализован в цитоплазме.
Помимо -кетоглутаровой кислоты, играющей основную роль в первичном связывании аммиака, роль акцепторов аммиака в растениях могут выполнять и другие органические кислоты, которые с помощью соответствующих ферментов взаимодействуют с NH3, образуя так называемые первичные аминокислоты. Они же служат акцептором аминогрупп в различных реакциях переаминирования. К числу этих органических кислот относятся щавелевоуксусная, пировиноградная, гидроксипировиноградная, глиоксиловая и другие, в процессе восстановительного аминирования которых получаются соответственно аспарагиновая кислота, аланин, серин, глицин.
Принято считать, что образование аспарагина преобладает в том случае, когда происходит распад белков в семенах. В клетках корня и листьев растущего растения идет, главным образом, образование глютамина. Таким образом, образование аспарагина - это путь обезвреживания аммиака, появляющегося при распаде белка - так называемая регрессивная ветвь азотного обмена, тогда как синтез глютамина - это путь обезвреживания аммиака при синтезе белка - прогрессивная ветвь азотного обмена.







