В возбуждающих синапсах нервной системы медиатором может являться ацетилхолин, норадреналин, дофамин, серотонин, глугаминовая кислота, вещество Р, а также большая группа других веществ, являющихся, если не медиаторами в прямом значении, то во всяком случае модуляторами (меняющими эффектиьность) синаптической передачи. Возбуждающие медиаторы вызывают появление на постсинаптической мембране возбуждающего постсинаптического потенциала (ВПСП). Его формирование обусловлено тем, что медиатор-рецепторный комплекс активирует Na- каналы мембраны (а также вероятно и Са-каналы) и вызывает за счет поступления натрия внутрь клетки деполяризацию мембраны. Одновременно происходит и уменьшение выхода из клетки ионов К+Амплитуда одиночного ВПСП однако довольно мала, и для уменьшения заряда мембраны до критического уровня деполяризации необходима одновременная активация нескольких возбуждающих синапсов. ВПСП, образуемые на постсинаптической мембране этих синапсов, способны суммироваться, т.е. усиливать друг друга, приводя к росту амплитуды ВПСП (пространственная суммация). Растет амплитуда ВПСП и при увеличении частоты поступающих к синапсу нервных импульсов (временная суммация), что повышает число выводимых в синаптическую щель квантов медиатора. Процесс спонтанной регенеративной деполяризации возникает в нейроне обычно в месте отхождения от тела клетки аксона, в так называемом аксонном холмике, где аксон еше не покрыт миелином и порог возбуждения наиболее низкий. Таким образом, ВПСП, возникающие в разных участках мембраны нейрона и на его дендритах, распространяются к аксонному холмику, где суммируются, деполяризуя мембрану до критического уровня и приводя к появлению потенциала действия. Тормозной постсинаптический потенциал (ТПСП)В тормозных синапсах обычно действуют другие, тормозные, медиаторы. Среди них хорошо изученными являются аминокислота глицин (тормозные синапсы спинного мозга), гамма-аминомасляная кислота (ГАМК) — тормозной медиатор в нейронах головного мозга. Вместе с тем, тормозной синапс может иметь тот же медиатор, что и возбуждающий, но иную природу рецепторов постсинаптической мембраны. Так, для ацетилхолина, биогенных аминов и аминокислот на постсинаптической мембране разных синапсов могут существовать как минимум два типа рецепторов, и, следовательно, разные медиатор-рецепторные комплексы способны вызывать различную реакцию хемочувствительных рецепторуправляемых каналов. Для тормозного эффекта такой реакцией может являться активация калиевых каналов, что вызывает увеличение выхода ионов калия наружу и гиперполяризацию мембраны. Аналогичный эффект во многих тормозных синапсах имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Возникающий при гиперполяризации сдвиг мембранного потенциала получил название тормозного постсинаптического потенциала (ТПСП). На рис.3.5 показаны отличительные черты ВПСП и ТПСП. Увеличение частоты нервных импульсов, приходящих к тормозному синапсу, также как и в возбуждающих синапсах, вызывает нарастание числа квантов тормозного медиатора, выделяющихся в синаптическую щель, что, соответственно, повышает амплитуду гиперполяризационного ТПСП. Вместе с тем, ТПСП не способен распространяться по мембране и существует только локально. В результате ТПСП уровень мембранного потенциала удаляется от критического уровня деполяризации и возбуждение становится либо вообще невозможным, либо для возбуждения требуется суммация значительно больших по амплитуде ВПСП, т.е. наличие значительно больших возбуждающих токов. При одновременной активации возбуждаюших и тормозных синапсов резко падает амплитуда ВПСП, так как деполяризующий поток ионов Na+компенсируется одновременным выходом ионов К+ в одних видах тормозных синапсов или входом ионов СГ в других, что называют шунтированием ВПСП.
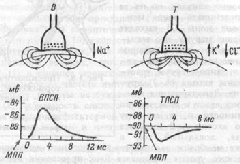 . МПП — мембранный потенциал покоя. Стрелки у синапсов показывают направление тока. Под влиянием некоторых ядов может происходить блокада тормозных синапсов в нервной системе, что вызывает безудержное возбуждение многочисленных рефлекторных аппаратов и проявляется в виде судорог. Так действует стрихнин, конкурентно связывающий рецепторы постсинаптической мембраны и не позволяющий им взаимодействовать с тормозным медиатором. Столбнячный токсин, нарушающий процесс освобождения тормозного медиатора, также угнетает тормозные синапсы. Принято различать два типа торможения в нервной системе: первичное и вторичное Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением: наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей. Этими особенностями являются следующие. 1. Иррадиация (дивергенция) возбуждения в ЦНС. Она объясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся (рис. 4.4, а). Иррадиацию возбуждения можно наблюдать в опыте на спинальной лягушке, когда слабое раздражение вызывает сгибание одной конечности, а сильное – энергичные движения всех конечностей и даже туловища. Дивергенция расширяет сферу действия каждого нейрона. Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов.
. МПП — мембранный потенциал покоя. Стрелки у синапсов показывают направление тока. Под влиянием некоторых ядов может происходить блокада тормозных синапсов в нервной системе, что вызывает безудержное возбуждение многочисленных рефлекторных аппаратов и проявляется в виде судорог. Так действует стрихнин, конкурентно связывающий рецепторы постсинаптической мембраны и не позволяющий им взаимодействовать с тормозным медиатором. Столбнячный токсин, нарушающий процесс освобождения тормозного медиатора, также угнетает тормозные синапсы. Принято различать два типа торможения в нервной системе: первичное и вторичное Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением: наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей. Этими особенностями являются следующие. 1. Иррадиация (дивергенция) возбуждения в ЦНС. Она объясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся (рис. 4.4, а). Иррадиацию возбуждения можно наблюдать в опыте на спинальной лягушке, когда слабое раздражение вызывает сгибание одной конечности, а сильное – энергичные движения всех конечностей и даже туловища. Дивергенция расширяет сферу действия каждого нейрона. Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов.
 Рис. 4.4. Дивергенция афферентных дорсальных корешков на спинальные нейроны, аксоны которых, в свою очередь, ветвятся, образуя многочисленные коллатерали (в), и конвергенция эфферентных путей от различных отделов ЦНС на α-мотонейрон спинного мозга (6) 1. Конвергенция возбуждения (принцип общего конечного пути) – схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу (принцип шеррингтоновской воронки). Конвергенция возбуждения объясняется наличием многих аксонных коллатералей, вставочных нейронов, а также тем, что афферентных путей в несколько раз больше, чем эфферентных нейронов. На одном нейроне ЦНС может располагаться до 10 000 синапсов. Явление конвергенции возбуждения в ЦНС имеет широкое распространение. Примером может служить конвергенция возбуждения на спинальном мотонейроне. Так, к одному и тому же спинальному мотонейрону подходят первичные афферентные волокна (рис. 4.4, б), а также различные нисходящие пути многих вышележащих центров ствола мозга и других отделов ЦНС. Явление конвергенции весьма важно: оно обеспечивает, например, участие одного мотонейрона в нескольких различных реакциях. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чиханья и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг. На рис. 4.4, я показаны два афферентных волокна, каждое из которых отдает коллатерали к 4 нейронам таким образом, что 3 нейрона из общего их числа, равного 5, образуют связи с обоими афферентными волокнами. На каждом из этих 3 нейронов конвергируют два афферентных волокна. На один мотонейрон может конвергировать множество коллатералей аксонов, до 10 000-20 000, поэтому генерация ПД в каждый момент зависит от общей суммы возбуждающих и тормозящих синаптических влияний. ПД возникают лишь в том случае, если преобладают возбуждающие влияния. Конвергенция может облегчать процесс возникновения возбуждения на общих нейронах в результате пространственной суммации подпороговых ВПСП либо блокировать его вследствие преобладания тормозных влияний (см. раздел 4.8). 3. Циркуляция возбуждения по замкнутым нейронным цепям. Она может продолжаться минуты и даже часы (рис. 4.5).
Рис. 4.4. Дивергенция афферентных дорсальных корешков на спинальные нейроны, аксоны которых, в свою очередь, ветвятся, образуя многочисленные коллатерали (в), и конвергенция эфферентных путей от различных отделов ЦНС на α-мотонейрон спинного мозга (6) 1. Конвергенция возбуждения (принцип общего конечного пути) – схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу (принцип шеррингтоновской воронки). Конвергенция возбуждения объясняется наличием многих аксонных коллатералей, вставочных нейронов, а также тем, что афферентных путей в несколько раз больше, чем эфферентных нейронов. На одном нейроне ЦНС может располагаться до 10 000 синапсов. Явление конвергенции возбуждения в ЦНС имеет широкое распространение. Примером может служить конвергенция возбуждения на спинальном мотонейроне. Так, к одному и тому же спинальному мотонейрону подходят первичные афферентные волокна (рис. 4.4, б), а также различные нисходящие пути многих вышележащих центров ствола мозга и других отделов ЦНС. Явление конвергенции весьма важно: оно обеспечивает, например, участие одного мотонейрона в нескольких различных реакциях. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чиханья и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг. На рис. 4.4, я показаны два афферентных волокна, каждое из которых отдает коллатерали к 4 нейронам таким образом, что 3 нейрона из общего их числа, равного 5, образуют связи с обоими афферентными волокнами. На каждом из этих 3 нейронов конвергируют два афферентных волокна. На один мотонейрон может конвергировать множество коллатералей аксонов, до 10 000-20 000, поэтому генерация ПД в каждый момент зависит от общей суммы возбуждающих и тормозящих синаптических влияний. ПД возникают лишь в том случае, если преобладают возбуждающие влияния. Конвергенция может облегчать процесс возникновения возбуждения на общих нейронах в результате пространственной суммации подпороговых ВПСП либо блокировать его вследствие преобладания тормозных влияний (см. раздел 4.8). 3. Циркуляция возбуждения по замкнутым нейронным цепям. Она может продолжаться минуты и даже часы (рис. 4.5).
 Рис. 4.5. Циркуляция возбуждения в замкнутых нейронных цепях по Лоренто де-Но (а) и по И.С.Беритову (б). 1,2,3- возбуждающие нейроны Циркуляция возбуждения – одна из причин явления последействия, которое будет рассмотрено далее (см. раздел 4.7). Считают, что циркуляция возбуждения в замкнутых нейронных цепях – наиболее вероятный механизм феномена кратковременной памяти (см. раздел 6.6). Циркуляция возбуждения возможна в цепи нейронов и в пределах одного нейрона в результате контактов разветвлений его аксона с собственными дендритами и телом. 4. Одностороннее распространение возбуждения в нейронных цепях, рефлекторных дугах. Распространение возбуждения от аксона одного нейрона к телу или дендритам другого нейрона, но не обратно объясняется свойствами химических синапсов, которые проводят возбуждение только в одном направлении (см. раздел 4.3.3). 5. Замедленное распространение возбуждения в ЦНС по сравнению с его распространением по нервному волокну объясняется наличием на путях распространения возбуждения множества химических синапсов. Время проведения возбуждения через синапс затрачивается на выделение медиатора в синаптическую щель, распространение его до постсинаптической мембраны, возникновение ВПСП и, наконец, ПД. Суммарная задержка передачи возбуждения в синапсе достигает примерно 2 мс. Чем больше синапсов в нейрональной цепочке, тем меньше общая скорость распространения по ней возбуждения. По латентному времени рефлекса, точнее по центральному времени рефлекса, можно ориентировочно рассчитать число нейронов той или иной рефлекторной дуги. 6. Распространение возбуждения в ЦНС легко блокируется определенными фармакологическими препаратами, что находит широкое применение в клинической практике. В физиологических условиях ограничения распространения возбуждения по ЦНС связаны с включением нейрофизиологических механизмов торможения нейронов. Рассмотренные особенности распространения возбуждения дают возможность подойти к пониманию свойств нервных центров.
Рис. 4.5. Циркуляция возбуждения в замкнутых нейронных цепях по Лоренто де-Но (а) и по И.С.Беритову (б). 1,2,3- возбуждающие нейроны Циркуляция возбуждения – одна из причин явления последействия, которое будет рассмотрено далее (см. раздел 4.7). Считают, что циркуляция возбуждения в замкнутых нейронных цепях – наиболее вероятный механизм феномена кратковременной памяти (см. раздел 6.6). Циркуляция возбуждения возможна в цепи нейронов и в пределах одного нейрона в результате контактов разветвлений его аксона с собственными дендритами и телом. 4. Одностороннее распространение возбуждения в нейронных цепях, рефлекторных дугах. Распространение возбуждения от аксона одного нейрона к телу или дендритам другого нейрона, но не обратно объясняется свойствами химических синапсов, которые проводят возбуждение только в одном направлении (см. раздел 4.3.3). 5. Замедленное распространение возбуждения в ЦНС по сравнению с его распространением по нервному волокну объясняется наличием на путях распространения возбуждения множества химических синапсов. Время проведения возбуждения через синапс затрачивается на выделение медиатора в синаптическую щель, распространение его до постсинаптической мембраны, возникновение ВПСП и, наконец, ПД. Суммарная задержка передачи возбуждения в синапсе достигает примерно 2 мс. Чем больше синапсов в нейрональной цепочке, тем меньше общая скорость распространения по ней возбуждения. По латентному времени рефлекса, точнее по центральному времени рефлекса, можно ориентировочно рассчитать число нейронов той или иной рефлекторной дуги. 6. Распространение возбуждения в ЦНС легко блокируется определенными фармакологическими препаратами, что находит широкое применение в клинической практике. В физиологических условиях ограничения распространения возбуждения по ЦНС связаны с включением нейрофизиологических механизмов торможения нейронов. Рассмотренные особенности распространения возбуждения дают возможность подойти к пониманию свойств нервных центров.








