В предыдущих главах было показано, что экстрагенитальные циклы у самок полициклических видов, суммируясь, определяют динамику их общей реактивности по фазам полового цикла. Для проверки и уточнения этого положения в качестве объекта изучения были выбраны различия радиочувствительности, т. е. реакции организма на действие ионизирующей радиации. Известно, что радиочувствительность отражает общую реактивность и может служить мерой резистентности организма (Русин, 1971).
В радиобиологии вопрос о половых различиях радиочувствительности остается неясным. На экспериментальном материале одни авторы делают заключение о более высокой радиочувствительности женских особей, другие считают более радиочувствительным мужской организм.
Есть работы, в которых показано отсутствие различий радиочувствительности в связи с полом.
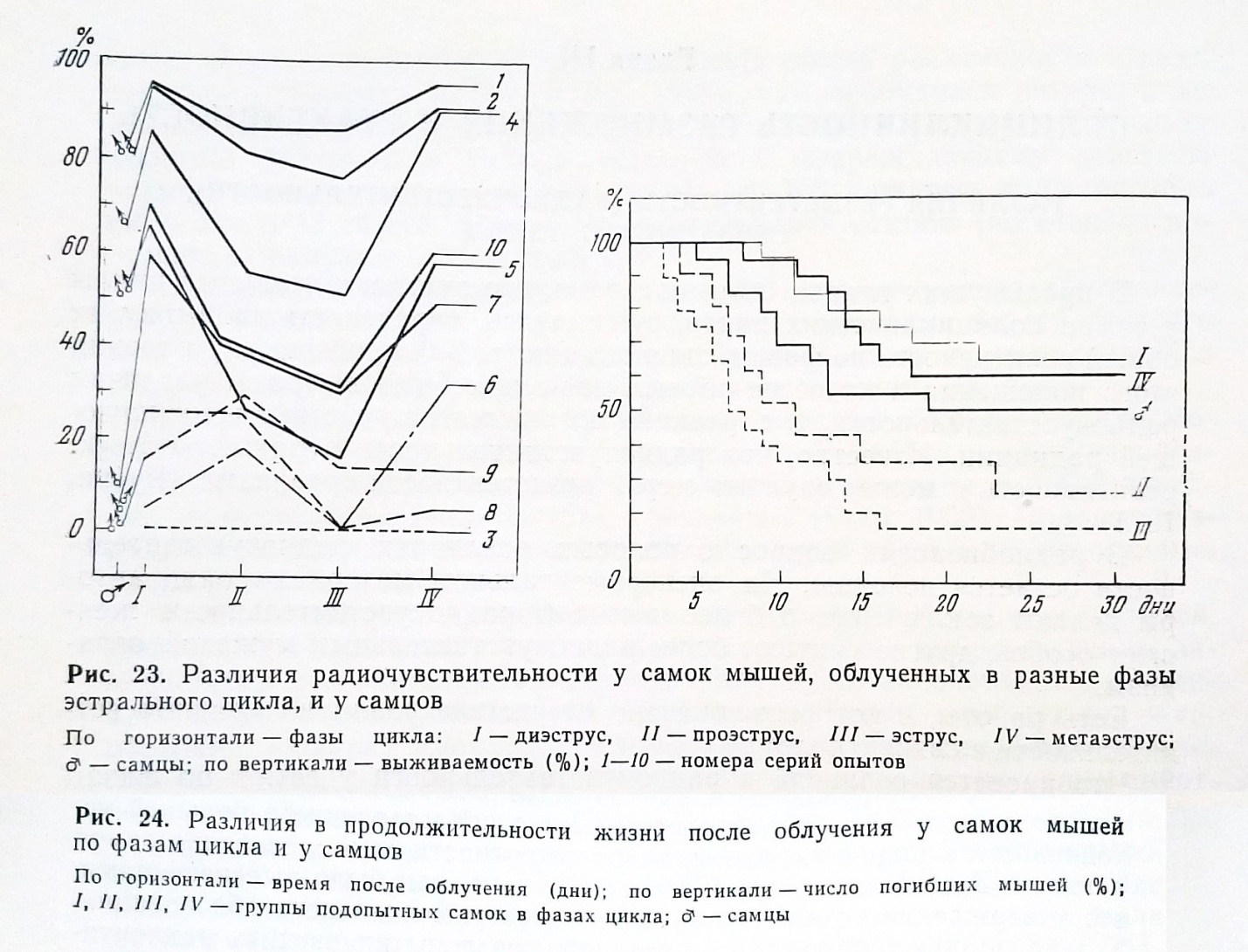
Что касается различий в радиочувствительности у самок по фазам полового цикла - в радиобиологию вошло представление о большей устойчивости к радиации, т. е. радиорезистентности самок, особенно самок в течке (Бак, Александер, 1963; и др.), хотя, как было выяснено нами, это утверждение не имело под собою экспериментального обоснования. Только в двух работах были предприняты попытки изучить радиочувствительность самок в разных фазах цикла. Rugh и Clugst'on (1955) в одной серии опытов получили большую устойчивость самок в фазе течки, но допустили методическую ошибку и не воспроизвели результата. Одну серию опытов с облучением самок по всем фазам цикла (в диэструсе, проэструсе, эструсе и метаэструсе) выполнили Н. И. Шапиро с соавторами (1958) и сделали вывод об отсутствии различий в радиочувствительности по фазам цикла, посчитав случайным небольшое увеличение процента гибели самок, облученных в течке. Поскольку ясной картины не сложилось, мы для проверки, изменяется ли и как именно радиочувствительность самок в связи со сменой фаз половых циклов, облучали мышей в дозах вызывавших гибель от 12 до 100%. В 10 сериях облучалось по 100 белых беспородных мышей (по 20 самок в четырех фазах цикла и по 20 самцов). Данные сведены в рис. 23. Различия в радиочувствительности выступили совершенно отчетливо и в пределах доз ЛД12-88/30 были статистически достоверными. Радиочувствительность самок была высокой в проэструсе и эструсе и почти вдвое снижалась в метаэструсе и диэструсе т е самки оказались значительно и достоверно более устойчивыми к действию ионизирующих излучений в метаэструсе и диэструсе. Самцы по чувствительности к радиации заняли промежуточное положение - были более радиочувствительными, чем самки, в устойчивых фазах цикла но несколько более радиорезистентны, чем самки в фазах проэструса и эструса, в фазах высокой радиочувствительности.
Эти различия отчётливо выступили и при сравнении распределения гибели мышей в течение первого месяца после облучения (рис. 24). Как видно из рисунка, доза в 800 р (серия 5), при которой общий процент гибели для всех 100 облученных мышей составил 55, привела к гибели половины самцов, 73 – в проэструсе, 85 – в эструсе, но только 35 – в диэструсе и 40 – в эструсе. При этом раньше всего гибель начиналась среди самцов и в группах самок, облученных в радиочувствительных фазах – проэструсе и эструсе.
89

Дополнительно были поставлены эксперименты для оценки различий в общей резистентности мышей в разных фазах эстрального цикла по их выносливости к экстремальной нагрузке. Как и в опытах с действием ионизирующего излучения, лабораторных белых мышей собирали в группы по 10 голов в каждой фазе эстрального цикла, партиями пускали плавать в ванну до полного изнеможения и отмечали для каждой мыши время, когда она переставала плавать и опускалась на дно. Результаты объединены в рис. 25, из которого видно, что менее выносливыми оказы-
90
вались мыши-самки в фазах проэструса и эструса. Они первыми отказывались от борьбы за жизнь и прекращали плавать. Несколько более выносливыми были самцы, и еще длительнее держались на воде мыши в фазах диэструса и метаэструса.
Этими опытами еще раз было подкреплено представление о положительной корреляции между радиочувствительностью и общей реактивностью (Русин, 1971; и др.).
С достаточной степенью достоверности было показано, что по фазам полового цикла общая резистентность и радиочувствительность изменяются почти вдвое в пределах полулетальных доз и при экстремальных нагрузках.
Полученные нами достоверные данные противоречат представлениям радиобиологов. Они убедительно показывают, что самки более резистентны в фазах метаэструса и диэструса, а не в фазах активной секреции эстрогенов, п что самцы по радиочувствительности сближаются с самками в менее резистентных состояниях. В последнее время это положение находит подтверждение в радиологической клинической практике (Гуськова, Байсаголов, 1971). Что касается механизма формирования фазовых различий реактивности и радиорезистентности самок, можно думать, что он сводится к интеграции периодических, синхронных смене фаз половых циклов изменений в направленности, скорости, интенсивности энергетических, обменных, функциональных и морфологических процессов жизнедеятельности организма самок - к экстрагенитальной цикличности.
Динамика реактивности и жизненных функций женских особей с ее волнообразной ритмичностью резко отличает их от особей мужского пола, для организма которых пока еще не установлено сколько-нибудь похожих периодических колебаний жизнедеятельности, хотя имеются указания на 10-дневные колебания в секреции тестотерона и на 4-недельную периодичность в проявлении эпилепсии у мужчин.
Поскольку половая функция у всех млекопитающих носит цикличный характер (репродуктивный цикл, половые циклы и пр.), постольку для самок, в отличие от самцов, характерна фазовая волнообразная динамика жизнедеятельности н реактивности организма (экстрагенитальный ритм), которая особенно ярко выражена у полициклических видов грызунов и у приматов, включая человека.
Исследуя изменения радиочувствительности у лабораторных животных, мы встретились с достаточно неожиданным явлением, которое потребовало специального изучения.
Как оказалось, для выявления различий радиочувствительности по фазам астрального цикла необходимо было не только соблюдать одинаковые условия обследования и облучения животных, но и отбирать самок по фазам цикла в возможно более короткие сроки. Если же самки ожидали своей очереди 2-3 часа или облучались через 2-3 часа после отбора по фазам, различия в радиочувствительности элиминировались. Так, неудачей закончился опыт с облучением большой партии крыс-самок, которых сортировали по фазам на протяжении более чем 2-х часов до облучения.
Проверка показала, что уже через 1 час (а в отдельных случаях через 15-30 мин.!) самки мышей, крыс и морских свинок из проэструса и эструса переходят в метаэструс и диэструс. Для этого достаточно было извлечь их из клетки, взвесить, взять у них вагинальный мазок и снова посадить на место.
Повторное обследование проводилось через 1-3 часа после отбора по фазам, после мнимого облучения и облучения в полулетальных дозах (табл. 5).
Полученные в наших опытах данные труднообъяснимы с точки зрения традиционных представлений о последовательности фаз астральных
91
циклов. Известно, что половые циклы представляют собой цепь морфофизиологических изменений, индуцируемых половыми гормонами - эстрогенами и прогестероном, которые секретируются в яичниках созревающими фолликулами и развивающимся после овуляции желтым телом. Изменения в продолжительности и структуре циклов должны иметь в своей основе изменения функций гормонопродуцирующих образований яичников. Каждая фаза цикла в норме характеризуется продолжительностью, определяемой временем секреции основных половых гормонов. В норме для самок мышей проэструс продолжается 18 час эструс - около суток. Метаэструс и диэструс весьма варьируют по продолжительности, но длятся не меньше суток, а чаще они значительно более продолжительны. Для того, чтобы метаэструс или диэструс могли наступить у самок за 15-30 мин. вместо десятков часов, секреция фолликулов, по-видимому, должна в кратчайший срок пли прекращаться или заменяться секрецией прогестерона. «Выключение» циклов в ответ на изменения в содержании, па обстановку опытов можно отнести к проявлениям ориентировочно-оборонительной реакции. Более сильные воздействия выключают половые циклы па более длительные сроки (рис 26 На графиках отчетливо видны изменения астральных циклов у самок мышеи в ответ на различные воздействия - в дни привыкания к обстановке (ориентировочно-оборонительная реакция), в ответ на мнимое облучение (стрессовая реакция) и, наконец, на разные дозы ионизирующе-
92

го излучения (500 и 700 р). Отчетливо видно, чем сильнее воздействие, тем длительнее задержки выключенных циклов в фазах диэструса и метаэструса - в состояниях повышенной резистентности. Вероятно, прекращение секреции фолликулина, что приводит к выключению циклов, является следствием повышенной секреции тропных гормонов гипофиза — гормонов адаптации в ответ на стрессирующие и повреждающие воздействия при одновременном торможении секреции гонадотропинов.
С этой точки зрения становится понятным противоречивое действие одного из радиобиологических законов - закона Бергонье-Трибондо (1906), согласно которому быстрее и легче повреждаются молодые и пролиферирующие клетки. Это верно по отношению к семенникам, в которых радиация повреждает в первую очередь созревающие и делящиеся семенные клетки-сперматогонии, тогда как зрелые сперматозоиды сохраняют жизнеспособность и могут в течение некоторого времени после облучения обеспечить оплодотворение самок. При достаточно высоких дозах наступает частичная или полная стерильность, что находит практическое применение, например, в сельском хозяйстве в борьбе с насекомыми-вредителями.
Чтобы прекратить размножение вредителей, в места заражения выпускают стерилизованных в инсектариях самцов.
Но закон Бергонье-Трибондо оказывается недействительным по отношению к яичникам. Известно, что в ответ на повреждающие воздействия в яичниках первыми повреждаются внутренние и зернистые оболочки зрелых фолликулов и прекращается секреция фолликулина (Varteresz, 1966; Карапетян, Варданян, 1967; и др.). Значительно более устойчивыми оказываются растущие молодые фолликулы, примордиальные фолликулы и особенно желтые тела. Повреждение структуры и нарушение секреторной функции фолликулов непременно приводит к остановке, выключению половых циклов.
Это хорошо совпадает с данными ряда радиобиологов, которые отмечали нарушения астральных циклов после облучения в разных дозах у самок-крыс, особенно после облучения их в течке (Drips, Ford, 1932), у самок морских свинок (Gehther, 1934), у самок обезьян (Алексеева,, 1959; Диковенко, 1959), с нашими наблюдениями за половыми циклами у самок мышей, крыс и морских свинок после облучения их в сублетальных дозах.
Однако имеется несколько сообщений о том, что у самок животных,, подвергнутых облучению в дозах 400-500 р нарушения половых астральных циклов наступали не сразу, а через 1,5-2,5 месяца после воздействия (Geller, 1930; Kullender, 1953; Бетц, 1961). Наконец, Н. И. Шапиро с соавторами (1962) и другие исследователи отмечали отсутствие нарушений астральных циклов у самок даже после полулетальных доз ионизирующей радиации. Эта часть публикаций противоречит данным большинства авторов и не соответствует тому, что установлено в отношении характера и порядка развития поражений в яичниках под действием радиации.
Запаздывание и тем более отсутствие нарушений в течение половых циклов у самок после облучения дозами в сотни рентгенов представляется совсем непонятным, так как при таких дозах радиации наблюдаются повреждения зрелых, продуцирующих эстрогены фолликулов. Трудно представить, чтобы половые циклы могли продолжаться у самок в обычной для них последовательности фаз в отсутствии индуцирующего их фолликулина и при наличии структурных изменений в яичниках. Несомненно, половые циклы у самок должны нарушаться и прекращаться в самые короткие после облучения сроки.
В наших опытах было также установлено, что половые циклы у самок лабораторных животных изменяются по продолжительности и по структуре вплоть до полной остановки не только после облучения, но и
94
в ответ на перемены в условиях содержания, на незнакомую, непривычную обстановку опыта, на обследование (взятие из клетки, взвешивание, вагинальные лаважи), при которых морфологических нарушений в яичниках не происходит. В таких случаях изменения в течении половых циклов носят функциональный характер. По-видимому, они связаны с ориентировочно-оборонительной реакцией на обстановку и процедуру обследования, являются одним из компонентов ориентировочно-оборонительной реакции у самок.
Механизм изменений половой цикличности у самок в условиях, не вызывающих поражения фолликулов, можно представить себе как переключение гипофиза на активную секрецию тройных гормонов адаптации, что непременно сопровождается прекращением секреции тропных половых гормонов - гонадотропинов и влечет за собою торможение гормонообразующей функции яичников. Если в яичниках эстрогены не вырабатываются в нужных количествах для обеспечения созревания фолликула и яйцеклетки, то растущие фолликулы регрессируют и яичник возвращается к исходному состоянию относительного покоя. Соответственно и половые циклы приостанавливаются в фазе покоя - диэструсе. Если же прекращение секреции фолликулина произойдет в достаточно зрелом фолликуле, то может начать развиваться желтое тело и наступит в яичниках лютеиновая фаза, а в половом цикле установится метаэструс.
В обоих случаях организм самки перейдет на более резистентный уровень жизнедеятельности. В этом и заключен, по нашему мнению, биологический смысл и ценность «защитного» переключения половой цикличности.
Было показано, что переключение на более резистентные фазы полового и экстрагенитального циклов зависит от силы воздействия и индивидуальных особенностей реактивности самок.
Обобщая данные литературы и собственные, мы пришли к выводам, которые, как нам кажется, могут иметь более широкое биологическое значение. Это, прежде всего, то, что самки, в особенности на протяжении репродуктивного периода жизни, находятся в двух различных по реактивности состояниях: более и менее резистентном. У самцов фазовые колебания реактивности пока не обнаружены. По своей чувствительности к воздействиям (и радиочувствительности) самцы сближаются с самками в менее устойчивых (более радиочувствительных) состояниях. Устойчивость и адаптивные возможности самок по сравнению с самцами повышены также за счет способности самок в короткие сроки (минуты) переходить на более резистентный уровень жизнедеятельности.
Установленные выше особенности реактивности связаны с половыми и экстрагенитальными циклами и наиболее ярко выражены у полициклических видов животных.






