

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...
Топ:
Когда производится ограждение поезда, остановившегося на перегоне: Во всех случаях немедленно должно быть ограждено место препятствия для движения поездов на смежном пути двухпутного...
Теоретическая значимость работы: Описание теоретической значимости (ценности) результатов исследования должно присутствовать во введении...
Характеристика АТП и сварочно-жестяницкого участка: Транспорт в настоящее время является одной из важнейших отраслей народного...
Интересное:
Что нужно делать при лейкемии: Прежде всего, необходимо выяснить, не страдаете ли вы каким-либо душевным недугом...
Влияние предпринимательской среды на эффективное функционирование предприятия: Предпринимательская среда – это совокупность внешних и внутренних факторов, оказывающих влияние на функционирование фирмы...
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Дисциплины:
|
из
5.00
|
Заказать работу |
Содержание книги
Поиск на нашем сайте
|
|
|
|
Можно представить в виде схемы:
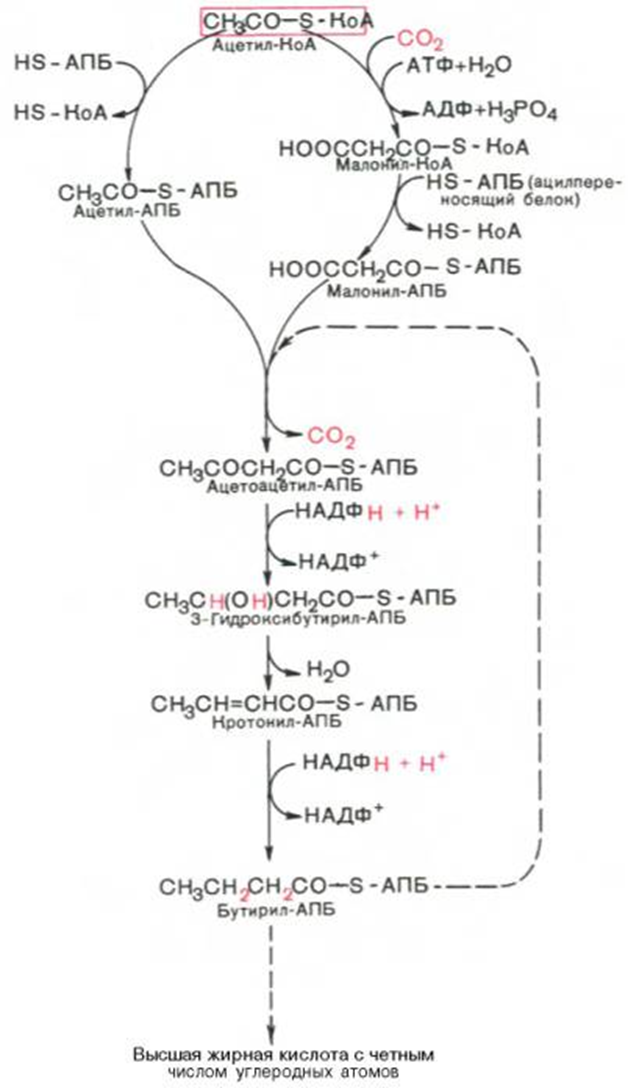
В общем виде синтез жирных кислот у кишечной палочки представлен на рис. 11.4. Последовательность и характер реакций в синтезе жирных кислот, начиная с образования β-кетоацил-АПБ (на рис. 11.4 – ацетоацетил-АПБ) и кончая завершением одного цикла удлинения цепи на два углеродных атома, являются как бы обратными реакциями окисления жирных кислот. На самом деле пути синтеза и окисления жирных кислот не пересекаются даже частично. Это становится очевидным, если принять во внимание некоторые особенности синтеза и окисления жирных кислот.
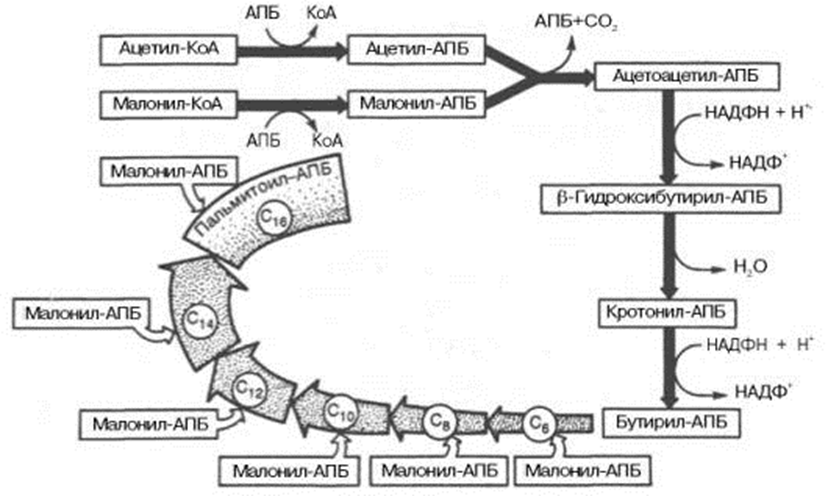
Синтез пальмитиновой кислоты у кишечной палочки при участии одной молекулы ацетил-КоА и 7 молекул малонил-КоА. Подробно представлен первый цикл синтеза - образование бутирил-АПБ. Остальные 6 циклов аналогичны первому.
По сравнению с β-окислением биосинтез жирных кислот имеет ряд характерных особенностей: синтез жирных кислот в основном осуществляется в цитозоле клетки, а окисление – в митохондриях; участие в процессе биосинтеза жирных кислот малонил-КоА, который образуется путем связывания СО2 (в присутствии биотин-фермента и АТФ) с ацетил-КоА; на всех этапах синтеза жирных кислот принимает участие ацилпереносящий белок (HS-АПБ); при биосинтезе образуется D(–)-изомер 3-гидроксикис-лоты, а не L(+)-изомер, как это имеет место при β-окислении жирных кислот; необходимость для синтеза жирных кислот кофермента НАДФН. Последний в организме частично (на 50%) образуется в реакциях пен-тозофосфатного цикла, частично – в других реакциях, в частности в реакциях:
Малат + НАДФ+-> Пируват + С02 + НАДФН + Н+ Изоцитрат + НАДФ+-> α-Кетоглутарат + С02 + НАДФН + Н +.
БИОСИНТЕЗ ТАГ
Известно, что скорость биосинтеза жирных кислот во многом определяется скоростью образования триглицеридов и фосфолипидов, так как свободные жирные кислоты присутствуют в тканях и плазме крови в небольших количествах и в норме не накапливаются.
Синтез триглицеридов происходит из глицерина и жирных кислот (главным образом стеариновой, пальмитиновой и олеиновой). Путь биосинтезатриглицеридов в тканях протекает через образование α-глице-рофосфата (глицерол-3-фосфата) как промежуточного соединения.
В почках, а также в стенке кишечника, где активность фермента глицеролкиназы высока, глицерин фосфорилируется за счет АТФ с образованием глицерол-3-фосфата: 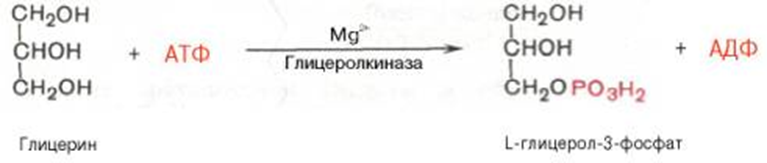
В жировой ткани и мышцах вследствие очень низкой активности глицеролкиназы образование глицерол-3-фосфата в основном связано с процессами гликолиза и гликогенолиза. Известно, что в процессе гли-колитического распада глюкозы образуется дигидроксиацетонфосфат (см. главу 10). Последний в присутствии цитоплазматической глицерол-3-фос-фатдегидрогеназы способен превращаться в глицерол-3-фосфат: 
Отмечено, что если содержание глюкозы в жировой ткани понижено (например, при голодании), то образуется лишь незначительное количество глицерол-3-фосфата и освободившиеся в ходе липолиза свободные жирные кислоты не могут быть использованы для ресинтеза триглицеридов, поэтому жирные кислоты покидают жировую ткань. Напротив, активация гликолиза в жировой ткани способствует накоплению в ней триглицеридов, а также входящих в их состав жирных кислот. В печени наблюдаются оба пути образования глицерол-3-фосфата.
Образовавшийся тем или иным путем глицерол-3-фосфат последовательно ацилируется двумя молекулами КоА-производного жирной кислоты (т.е. «активными» формами жирной кислоты – ацил-КоА). В результате образуется фосфатидная кислота (фосфатидат): 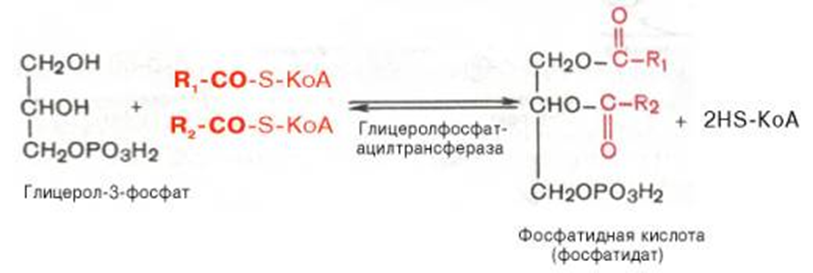
Как отмечалось, ацилирование глицерол-3-фосфата протекает последовательно, т.е. в 2 этапа. Сначала глицерол-3-фосфат-ацилтрансфераза катализирует образование лизофосфатидата (1-ацилглицерол-3-фосфата, а затем 1-ацилглицерол-3-фосфат-ацилтрансфераза катализирует образование фосфатидата (1,2-диацилглицерол-3-фосфата).
Далее фосфатидная кислота гидролизуется фосфатидат-фосфогидро-лазой до 1,2-диглицерида (1,2-диацилглицерола):

Затем 1,2-диглицерид ацилируется третьей молекулой ацил-КоА и превращается в триглицерид (триацилглицерол). Эта реакция катализируется диацилглицерол-ацилтрансферазой: 
Синтез триглицеридов (триацилглицеролов) в тканях происходит с учетом двух путей образования глицерол-3-фосфата и возможности синтеза триглицеридов в стенке тонкой кишки из β-моноглицеридов, поступающих из полости кишечника в больших количествах после расщепления пищевыхжиров. На рис. 11.6 представлены глицерофосфатный, дигидроксиацетон-фосфатный и β-моноглицеридный (моноацилглицероловый) пути синтеза триглицеридов.

Рис. 11.6. Биосинтез триглицеридов (триацилглицеролов).
Установлено, что большинство ферментов, участвующих в биосинтезе триглицеридов, находятся в эндоплазматическом ретикулуме, и только некоторые, например глицерол-3-фосфат-ацилтрансфераза,– в митохондриях.
Биосинтез триглицеридов (триацилглицеролов).
Биосинтез жирных кислот Жир может синтезироваться как из продуктов распада жира, так и из углеводов. Синтез эндогенного жира из углеводов протекает в печени и в жировой ткани. Ацил-КоА синтезируется из Ацетил-КоА. ГБФ-путь распада углеводов обеспечивает синтез энергией. Образование Ацетил-КоА происходит в матриксе митохондрий. Синтез жирных кислот идет в цитоплазме на мембранах эндоплазматического ретикулума путем постепенного удлинения ацетил-КоА на 2 углеродных атома за каждый цикл. Удлинение высших жирных кислот, содержащих более 16 углеродных атомов, идет путем реакций, обратных -окислению. Однако реакции синтеза жирных кислот до 16 углеродных атомов принципиально отличаются от реакций, обратных -окислению. Они протекают обходным обратным путем.
Гормональная регуляция
Липогенез – процесс синтеза жирных кислот, интенсивно протекающий в печени и жировой ткани. Следует отметить, что биосинтез липидов осуществляется в основном в цитоплазме клетки и лишь незначительная их часть, как считают некоторые авторы, образуется в митохондриях.
Образовавшийся в митохондриях ацетил-КоА способен проникать через их мембрану в цитоплазму только после взаимодействия с оксалоацетатом и образования цитрата, который достигает цитоплазмы, где вновь превращается в ацетил-КоА и оксалоацетат. Таким образом, оксалоацетат выполняет функцию катализатора, способствуя переносу ацетильной группы из интрамитохондриального пространства в цитозоль, где при участии ацетил-КоА-карбоксилазы происходит карбоксилирование ацетил-КоА с образованием малонил-КоА. Последний, взаимодействуя с комплексом жирные кислоты – белок при участии АТФ и КоА, превращается в комплекс жирнокислотный ацетил-КоА, что является необходимой стадией образования триглицеридов.
Карбоксилирование ацетил-КоА является важным регулятором синтеза жирных кислот и на всех последующих этапах липогенеза участвуют либо малонил-КоА, либо ацетил-КоА. Так под влиянием синтетазы жирных кислот и при использовании одной молекулы ацетил-КоА и 7 молекул малонил-КоА образуется пальмитил-КоА, способный к разнообразным превращениям. При потере одной молекулы СО2 и КоА образуется пальмитиновая кислота, молекула которой содержит 16 атомов углерода. Путем удлинения цепи в митохондриях или в плазматическом ретикулуме до 18 атомов углерода пальмитиновая кислота может быть превращена в стеариновую, а после десатурации она же преобразуется в пальмитоолеиновую и олеиновую кислоты.
При эстерификации жирных кислот с глицерином образуются триглицериды, причем в реакции участвует не свободный глицерин, а его производное – глицерол-3-фосфат, образующийся в печени из глицерина при участии глицеролкиназы. Эстерификация жирных кислот в жировой ткани может происходить лишь при достаточном поступлении дигидроксиацетонфосфата, который образуется в процессе гликолиза и конвертируется в глицерол-3-фосфат при участии глицерофосфатдегидрогеназы.
В печени и жировой ткани инсулин увеличивает синтез жирных кислот и триглицеридов. При эндогенном биосинтезе триглицеридов, кроме глюкозы, могут использоваться соединения, образующиеся из гликогенных аминокислот. Липогенез в печени контролируется тиротропным гормоном и гормонами щитовидной железы. Так, после гипофизэктомии синтез липидов печенью уменьшается и восстанавливается после введения тироксина.
основным местом образования плазменных пре-β-липопротеинов (липопротеины очень низкой плотности – ЛПОНП) и α-липопротеинов (липопротеины высокой плотности – ЛПВП) является печень.
образование ЛПОНП. Согласно данным литературы, основной белок апопротеин В-100 (апо Б-100) липопротеинов синтезируется врибосомах шероховатого эндоплазматического ретикулума гепатоцитов. В гладком эндоплазматическом ретикулуме, где синтезируются и липидные компоненты, происходит сборка ЛПОНП. Одним из основных стимулов образования ЛПОНП является повышение концентрации неэстерифици-рованных жирных кислот (НЭЖК). Последние либо поступают в печень с током крови, будучи связанными с альбумином, либо синтезируются непосредственно в печени. НЭЖК служат главным источником образования триглицеридов (ТГ). Информация о наличии НЭЖК и ТГ передается на мембранно-связанные рибосомы шероховатого эндоплазматического ретикулума, что в свою очередь является сигналом для синтеза белка (апо В-100). Синтезированный белок внедряется в мембрану шероховатого ретикулума, и после взаимодействия с фосфолипидным бислоем отмембраны отделяется участок, состоящий из фосфолипидов (ФЛ) и белка, который и является предшественником ЛП-частицы. Далее белокфосфо-липидный комплекс поступает в гладкий эндоплазматический ретикулум, где взаимодействует с ТГ и эстерифицированным холестерином (ЭХС), в результате чего после соответствующих структурных перестроек формируются насцентные, т.е. незавершенные, частицы (н-ЛПОНП). Последние поступают через тубулярную сеть аппарата Гольджи в секреторные везикулы и в их составе доставляются к поверхности клетки, после чего очень низкой плотности (ЛПОНП) в печеночной клетке (по А.Н. Климову и Н.Г. Никульчевой).
путем экзоцитоза выделяются в перисинусоидные пространства (пространства Диссе). Из последнего н-ЛПОНП поступают в просвет кровяного синусоида, где происходят перенос апопротеинов С из ЛПВП на н-ЛПОНП и достраивание последних (рис. 16.3). Установлено, что время синтеза апо В-100, образования липид-белковых комплексов и секреции готовых частиц ЛПОНП составляет 40 мин.
У человека основная масса β-липопротеинов (липопротеины низкой плотности – ЛПНП) образуется в плазме крови из ЛПОНП при действии липопротеинлипазы. В ходе этого процесса образуются сначала промежуточные короткоживущие липопротеины (Пр.ЛП), а затем формируются частицы, обедненные триглицеридами и обогащенные холестерином, т.е. ЛПНП.
При высоком содержании жирных кислот в плазме их поглощение печенью возрастает, усиливается синтез триглицеридов, а также окисление жирных кислот, что может привести к повышенному образованию кетоновых тел.
54. Липо́лиз.
Процесс расщепления жиров на составляющие их жирные кислоты под действием липазы.
Липолиз протекает в митохондриях, куда жирные кислоты доставляются с помощью переносчика — карнитина. В процессе липолиза происходят циклические превращения молекул жирных кислот с отщеплением от них двухуглеродных производных KoA (CH3—CO—SKoA) (β-окисление жирных кислот) или одноуглеродных производных KoA (α-окисление жирных кислот); протекание одного цикла окисления жирной кислоты, сопровождается синтезом по одной молекуле ФАДН и НАДН.
Фермент триглицеридлипаза расщепляет триглицериды на диглицериды и жирные кислоты, на следующей стадии активны диглицеридлипаза и моноглицеридлипаза. В результате работы этих ферментов образуются конечные продукты липолиза — глицерин и жирные кислоты.
Липолиз является важнейшим энергетическим процессом в клетке, который обеспечивает синтез самого большого количества АТФ. Например, при окислении одной молекулыпальмитата (CH3(CH2)14COOH), образуется 131 молекула АТФ, две из которых используются для активации пальмитата.
|
|
|

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...

История развития пистолетов-пулеметов: Предпосылкой для возникновения пистолетов-пулеметов послужила давняя тенденция тяготения винтовок...

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Историки об Елизавете Петровне: Елизавета попала между двумя встречными культурными течениями, воспитывалась среди новых европейских веяний и преданий...
© cyberpedia.su 2017-2026 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!

