Биологическое окисление.
Основным путем выделения энергии служит окислительное расщепление Именно окислительным путем разрывается основное количество химических святей питательных веществ Например аминокислоты образующиеся при гидролизе белка расщепляются затем до углекислого газа воды и аммиака Какие же способы окисления веществ реализуются в клетке?
Соединения могут окислятся по современным представлениям тремя различными способами
1 Реакции оксиденации Т е окисление путем прямого присоединения кислорода к окисляемому субстрату 2 Путь передачи электронов Некоторые реакции окислительные сопровождаются только потерей электронов, а затем происходит присоединение протонов 3 главный (основной) Путем отщепления атомов водорода от окисляемого субстрата - дегидрирование Принято различать 2 вида дегидрирования
а) Аэробное б) Анаэробное
В чем различие? Суть различия сводится к вопросу о первичном акцепторе отщепленного водорода Если атомы водорода отщепленные от субстрата переносятся сразу на кислород - аэробное Если отщепленные атомы водорода переносятся на соединения отличные от кислорода - анаэробное
Классификация фермента» участвующих в биоокислении. Оксиредуктазы Все ферменты катализирующие окислительно-восстановительные процессы Какие же здесь группы
1 Отщепление атомов водорода от окисляемого субстрата катализируется ферментами оегиАрогинюами Их разделяют на 2 подподкласса
а) Аэробные дегидрогиназы
б) Анаэробные дегидрогиназы
В чем разница между ними? Аэробные дегидропшазы катализируют перенос отщепленных атомов водорода от окисляемого субстрата на кислород в итоге образуется токсичная перекись водорода
H-S-H + О2 -> S окисл + Н2О2
Анаэробные дегидрогиназы катализируют перенос отщепленных атомов водорода на какое-то соединение отличающееся от кислорода (НАД, ФАД, ФМН), а субстрат окисляется, потеряв 2 атома водорода
H-S-H + X ->ХН2 + Зокисл
2 Оксигеназы Ферменты катализирующие присоединение кислорода или реакцию оксидинации Принято длить на 2 группы
а) Монооксигеназы (гидроксилазы) присоединяют атомарный кислород к окисляемому субстрату
б) Дноксигеназы присоединяют молекулу кислорода а) р
H-S + О2 + КН2 -» S ^H2O + Кокисл
Реакции монооксигеназного типа требуют еще одного участника так называемого Косубстрата Чаще всего выступает восстановленный НАД К субстрату присоединяется один атом кислорода
б) H-S-H + О2 >HO- S-OH ->S--O + H2O
3 Цитохромы Катализируют окисление веществ путем отдачи электронов Гемовое железо В одном из цитохромов имеется так же атом меди.
4 Вспомогательные ферменты биологического окисления К ним относятся такие ферменты как католаза и пероксидаза. которые играют защитную роль разрушая перекись водорода или органические перекиси образующихся в ходе окислительных процессах Перекиси представляют собой достаточно агрессивные соединения которые могут вызвать значительные изменения в клеточных структурах ФУНКЦИИ БИОЛОГИЧЕСКОГО ОКИСЛЕНИЯ
1 Важнейшей функцией биологического окисления является несомненно высвобождение энергии которая в дальнейшем используется в эндоорганических процессах
2 В ходе окисления питательных веществ образуется ряд низкомолекулярных соединений, которые клетка использует потом для биосинтеза Назыв - пластическая функция Например синтез аминокислот из продуктов окисления глюкозы или жиров
используется в биосинтетических реакциях Генерация восстановительных эквивалентов (потенциалов). 4
Окислительные процессы несут защитную роль Многие ксенобиотики обезвреживаются путем окисления в том числе многие лекарственные препараты 5. Огромная роль в поддержании температуры тела Таким образом существование живых существ невозможно без окислительных процессов
Цикл кребса.
Это высокоорганизованная циклическая система взаимопревращений ди- и трикарбоновых кислот, катализируемых мультиферментным комплексом Он составляет основу клеточного метаболизма Несмотря на то, что этот метаболический путь является замкнутым его началом считается цитратсинтазная реакция в ходе которой конденсация Ацетия-КоА и оксалоацитата дает цитрат Далее следует реакция отщепление воды катализируемая ферментом аконитазой продуктом реакции является цис- Этот же фермент (аконитаза или аконитатгитротаза) катализирует реакцию гидротации в итоге образуется изомер -изолимонная кислота Окислительная реакция кот катализируется ферментом изоцитратдегидрогиназа (она еще и карбоксилирующая) а-кетоггаотаровая кислота В ходе реакции отщепляется СО2, энергия окислительного превращения аккумулируется в восстановленном НАД а-кетоглютаровая кислота под действием а-кетоглюторатдегидрогиназного комплекса (3 типа ферментов, 2 кофермента НАД, КоА и 3 простетических группы тиаминпирофосфат, липоевую кислоту и Флавинадениндннуклеотид) Это тоже реакция окисления идущая путем дегидрирования В данном случае часть энергии запасается еще и в макроэргической связи Образуется сукценил-КоА Сукцешга-КоА Фермент катализирует очень интересную реакцию в ходе которой из ГДФ и фосфорной кислоты образуется ГТФ Отщепляется КоА фермент - сукценил-КоА-синтаза (сукцинаттиокиназа) В итоге образуется янтарная кислота - сукцинат Эта реакция субстратно-окислительного фосфорноирования Сукцинат далее вступает вновь в реакцию окисления с участием фермент» сукцинатдегидрогиназы Это ФАД зависимый фермент остальные дегидрогиназы НАД зависимые ФАД восстанавливается а сукцинат окисляется с образованием фумаровой кислоты Происходит немедленное присоединение воды с участием фермента фумаразы и образуется малат (яблочная кислота) Малат, с участием малатдегидрогиназы содержащий НАД, окисляется в итоге образуется ЩУК т е происходит регенерация первого продукта ЩУК может снова вступать в реакцию конденсации с ацетил-КоА с образованием лимонной
Что происходит в цикле Кребса? СНЗ-С + ЗНАД + ФАД + ГДФ + НЗРО4 + 2Н2О -> 2СОг + ЗНАДН+Н* + ФАДН2 + ГТФ + HSKoA
Как регулируется цикл Кребса? Термодинамический контроль работы цикла
Пусковой реакцией цикла является цитратсиктазная реакция она сопровождается большой потерей энергии Д G = - 7,7 ккал/моль В условиях клетки эта реакция практически необратима Большой потерей энергии сопровождается <х-кетоглюторатдегидрогиназная реакция ДО = -8 ккал/моль В условиях клетки эта реакция абсолютна необратима За счет пусковой стадии и особенно а-кетст люторатдегидрогиназной реакции движение метаболитов возможно лишь в одном направлении т е в направлении деградации (ращипления) ацетип-КоА т е здесь мы имеем дело с двойным термодинамическим контролем направления процесса Кинетический контроль скорости потока метаболитов. Обеспеченность клетки энергией можно характеризовать величиной энергетического ее заряда Он равен отношению концентрации АТФ и половины концентрации АДФ к концентрация АДФ, АТФ и АМФ Когда клетка хорошо обеспечена энергией большая часть ее адениловых нуклеотидов находится в форме АТФ и величина энергетического заряда приближается к 1 Высокая концентрация АДФ и АМФ признак нехватки энергии Отсюда работа цикла Кребса должна тормозиться высокой концентрацией АТФ Цикл Кребса должен стимулироваться высокой концентрацией АДФ и АМФ, энергии не хватает - пора включать цикл Аналогичную роль может играть отношение восстановленно! о НАД к окисленному Скорость работы цикла контролируется прежде всего на пусковой стадии, которая определяется в значительной мере концентрацией ЩУК в клетке Кроме того цитратсинтаза аллостерически угентается высокими концентрациями АТФ, восстав НАД, и сукцинил-КоА Угнетение активности ферментов высокими концентрациями АТФ вполне понятно - клетка обеспеченна энергией и цикл Кребса выключается Второе регуляторное звено - изоцитратдегидрогиназная реакция Изоцитратдегндрогиназа аллостерически ингибируется высокими концентрациями АТФ и восстановленного НАД В то же время АДФ выступает в качестве аллостерического активатора Аллостерическим активатором по некоторым данным является так же изоцитрат
Третьим уровнем регуляции является а-кетоглюторатдегидропшазная реакция Здесь восстановленный НАД и АТФ явл аллостер ингибиторами работы этого комплекса Активатором выступает АМФ Высокие концентрации сукцинил-КоА угнетает работу комплекса
Активность фермента сукцинатдегидрогиназы ингибируется по конкурентному механизму высокими концентрациями ЩУК и малоновой кислоты.
Классификация липидов.
Липиды - природные органические соединения (крайне гегерогенны по своей химической структуре) общими свойствами которых является низкая растворимость в воде и хорошая растворимость в аполярных растворителях таких как хлороформ, жидкие углеводороды и др
/. Жирные кислоты и их производные. - это алифатические карбоновые кислоты число атомов в которых может достигать 22-24 Они подразделяются на насыщенные жирные кислоты - не имеющие в своей структуре двойных связей И ненасыщенные жирные кислоты - имеющие в своей структуре двойные или даже тройные С - С связи (тройные встречаются крайне редко)
Ненасыщенные жирные кислоты в свою очередь делятся на
а) моноеновые те содержащие одну двойную связь
б) полиеновые, содержащие много двойных связей (диеновые, триеновые и др)
Природные ненасыщенные жирные кислоты (незаменимые) обычно имеют тривиальное название, например алеиновая, линоливая, линоленовая арахндоновая
Жирные кислоты в организме выполняют несколько функций. Прежде всею несомненно это энергетическая функция. Так же выполняют структурную функцию. Выполняют пластическую функцию Из ацетилКоА (продукт распада жирных кислот) в гепатоцитах синтезируются ацетоновые тела, холестнрол А эйкоюполяевовые кислоты используются для синтеза рядя биорегуляторов это простогландины. тромбоксаны, Особенно необходимо подчеркнуть, что ряд полиненасьпценных жирных кислот относятся к незаменимымВажную роль в регуляции функционирования клеток различных органов и тканей играет производные эйкозопояиеновых кислот, так называемые эйкозоноиды. К ним относятся простоноиды а) простогландины, 6) простоциклины, ъ)лейкотриены; г) трамбоксаны Первые три группы соединений (простогландины, простошклины, лейкотриены) объединяют так же в группу простоноиды Эйкозополиеновые кислоты - высшие жирные кислоты с 20 атомами углерода в цепи и имеющие в своей структуре несколько двойных связей
Простогландины, которые делятся на простогландины а, в, с, d и т д относятся к виорегуляторам паракринной системы. При очень низких концентрациях они вызывают сокращение гладкой мускулатуры, 1 участвуют в развитии воспалительной реакции. 2 они принимают участие в регуляции процесса свертывания крови, и 3 регулируют метаболические пути на уровне клеток Иначе их называют местными гормонами Тромбоксаны образуются в тромбоцитах и после выхода в кровяное русло вызывают сужение кровеносных сосудов и агрегацию тромбоцитовПростоцикянни образуются в стенках кровеносных сосудов и являются сильными ингибиторами агрегации тромбоцитов Лейкотриены представляют собой группу триенов с сопряженными двойными связями Они образуются в тромбоцитах, лейкоцитах и макрофагах в ответ на имуниологические и неимуннологические стимулы а) принимают участие в развитии анофелоксии, б) повышают проницаемость кровеносных сосудов, в) вызывают приток и активацию лейкоцитов Я. Глициринсодержащие липиды.Из глициринсодержащих липидов наибольшее значение имеют и Обычно их рассматривают как производные трехатомного спирта – глицерола делятся по количеству входящих в их состав ацильиых групп на а) моноацилглицигины -1 жирный кислотный остаток б)диадилпшцериныв)триацилглицерины Триайилглицерины. составляют основную массу резервных липидов человеческого организма Триацилглицерины выполняют резервную функцию Причем это преимущественно энергетический резерв организма Глицерол, входящий в структуру триацилглицерннов, может использоваться для синтеза глюкозы или некоторых2) Являясь одним из основных компонентов жировой ткани, триацилглицерины участвуют в защите внутренних органов человека от механических повреждений 3) Участвуют в терморегулящии, образуя теплоизолирующую прослойку Все глицерофосфолипиды можно рассматривать как производные фосфотидной кислоты которой один атом заменен на аминоспирт Основной функцией глицерофосфолипидов является структурная Они входят в качестве важнейших структурных компонентов в состав мембран 2)Некоторые гпицерофосфолнпиды выполняют специфические функции Например инозитолфосфотиды участвуют в работе регуляторных механизмов в клетке Ш. Липиды, не содержании глицерола.К этим липидам относятся множество самых разнообразных соединений химической природы Мы остановимся только на трех группах веществ имеющих высокую биологическую значимость а) сфинголипиды б) стероиды в) полипреноиды Сфинголипиды. Можно рассматривать как производные стерамида Отдельные классы сфингошгшдов отличаются друг от друга только характером группировки присоединенной функния сфшхалипидов Прежде всего структурная функция Они входят обязательно в состав клеточных мембран Углеводные компоненты цереброзидов и ганпшозидов участвуют в образовании гдикокаликса Причем в этом качестве они играют определенную роль во - первых в реализации межклеточных взаимодействий во - вторых во взаимодействии клеток с компонентами межклеточного вещества3) Ганглиозиды выполняют рецепторные функцииСтероиды. К ним относятся соединения имеющие в своей структуре стерановое ядро Различные соединения из класса стероидов отличаются друг от другаа) дополнительными углеводородными радикалами, б) наличием двойных связей,в) наличием различных функциональных групп г) различия могут ноешь стереохимический характер Биологически важные соединения сгпероидной природы
1) Холистерол 2) Стероидные гормоны (гормоны коры надпочечников глюко- и минералокортикоиды) 3)
Половые гормоны (андрогены и эстрогены)
СТЕРИДЫ И СТЕРИНЫ
Стернды представляют весьма важную группу соединений липоидного характера В отличие от фосфатидов они являются сложными эфирами жирных кислот и стеринов
С теряны— это циклические спирты, производные циклопентанопергидрофенантрена
Циклопентанопергидрофенантрен можно рассматривать как продует конденсации циклопентана и полностью гидрированного фенантрена (пергидрофенантрена)
Стерины широко распространены в растительном и животном мире В бактериях их обычно обнаружить не удается Стерины находятся в биологических объектах либо в свободном виде, либо в виде сложных эфиров с
жирными кислотами. Только 10% всех стеринов человеческого организма находятся в виде эфиров. Однако в
некоторых органах, например в печени, почти половина стеринов присутствует в форме сложных эфиров Стерины коровьего масла сала, а также стерины присутствующие в эритроцитах и желчи человека, находятся целиком в свободном состоянии
Холестерин. Изотдельных стеринов наибольший интерес для медиков представляют холестерин-
вторичный циклический спирт и его сложные эфиры — х о лестерид ы.
ОБмен холистерола.
Суточная потребность человека в холистероле составляет около 1 гр Причем вся потребность в этом соединении может удовлетворяться с помощью эндогенного синтеза Пищевой холистерол так же эффективно усваивается человеком > здорового человека поступление холистерола с пищей и его эндогенный синтез хорошо сбалансирован Так например поступление с пищей в течении суток 2-3 гр холистерола полностью блокирует его эндогенный синтез
Основным органом, в котором идет синтез холистерола является печень. В печени человека синтезируется от 50 до
80% эндогенного холистерола, 10-15% синтезируется в клетках тонкого кишечника и около 5% образуется в коже остальное в других органах и тканях Т е объем синтеза в других органах и тканях не названных (дентине, цементе) вообще незначителен, хотя ферментная система обеспечивающая синтез этого соединения присутствует практически во всех органах и тканях
В условиях обычного пищевого рациона во внутреннюю среду организма поступает около 300 мг экзогенного
Общее содержание холистерола в организме человека составляет примерно 140гр Основная масса этого соединения включена в состав клеточных мембран, однако около 10гр холистерола постоянно содержится в плазме крови, входя в состав липопротеидов Концентрация холистерола в норме составляет 3,5-6,8 млмоль/л Причем примерно всего 2/3 холистерола плазмы крови представлена в ней в виде сложных эфиров холистерина с жирными кислотами т е стероиды Жирные кислоты связанные с холистерином это преимущественно лгаюливая и олеиновая Избыток холистерола в клетках запасается в виде эфиров олеиновой кислоты, в то же время в состав мембран входит только свободный хотистерол.
Биологическая роль холестерола.
Холистерол используется в организме прежде всего
1 для синтеза желчных кислот в печени 2 из него синтезируются все стероидные гормоны 3 в коже из него образуется ^-дегидрохолистерин, который под действием УФ превращается в витамин D.
Обмен холестерола.
Суточная потребность человека в холистероле составляет около 1 гр Причем вся потребность в этом соединении может удовлетворяться с помощью эндогенного синтеза Пищевой холистерол так же эффективно усваивается человеком V здорового человека поступление \одистерола с пищей и его эндогенный синтез хорошо сбалансирован Так например поступление с пищей в течении суток 2-3 гр холистерола полностью блокирует его эндогенный синтез
Основным органом в котором идет синтез холистерола является в печень.В печени человека синтезируется от 50 до 80% эндогенного холистерола, 10-15% синтезируется в клетках тонкого кишечника и около 5% образуется в коже, остальное в других органах и тканях Т е объем синтеза в других органах и тканях не названных (дентине, цементе) вообще незначителен, хотя ферментная система обеспечивающая синтез этого соединения присутствует практически во всех органах и тканях. В условиях обычного пищевого рациона во внутреннюю среду организма поступает около 300 мг экзогенного хопистерола 500 - 700 мг холистерола организм обычно при смешанной диете получает за счет эндогенного синтеза. Общее содержание холестерола в организме человека примерно 140 гр. Основная масса этого соединения включена в состав клеточных мембран, однако около 10гр холистерола постоянно содержится в плазме крови, входя в состав липопротеидов Концентрация холистерола в норме составляет 3,5-6,8 млмоль/л Причем примерно всего 2/3 холистерола плазмы крови представлена в ней в виде сложных эфиров холистерина с жирными кислотами те стероиды. Жирные кислоты связанные с холистерином это преимущественно линоливая и олеиновая Избыток холистерола в клетках запасается в виде эфиров олеиновой кислоты, в то же время в состав мембран входит только свободный холистерол
Биологическая роль холистерола. Холистерол используется в организме прежде всего 1 для синтеза желчных кислот в печени 2 из на о синтезируются все стероидные гормоны 3 в коже из него образуется "'-дегидрохолистерин, который под действием УФ превращается в витамин D Как выводится холистерол? Избыток холистерола выводится из организма желчью. Последнее время доказано, что часть избыточного холнстерина может поступать в просвет кишечника непосредственно через его стенки Таким образом холистериновый гамеостаз в организме является результатом динамического равновесия во-первых процессов его поступления в организме эндогенного синтеза, и во-вторых процесов использования холистерола для нужд клеток и его выведение из организма Как синтезируется холистерол? Он синтезируется в клетках из двух углеродных группировок ацетилКоА Процесс синтеза включает в себя Зэ последовательных реакций и может быть разбит на 4-5 этапов
1 этап - образование из ацетилКоА мевалоновой кислоты
2 этап - -образование из меваяоновой кислоты активированных 5 углеродных группировок, гаопреноидные группировки (это изопентилпирофосфат, диметилаланиллипофосфат - активные изопеноидные группировки) 3 этап - конденсация изопреноидных группировок с образованием сквапена 4 этап - циклизация сквапена в ланоетерин 5 этап - преобразование ланостерина в холнстерол В ходе 1-й реакции которую катализирует ацетилКоА-адетилтрансфераза образуется ацетоацетилКоА Затем используется еще одна молекула ацетилКоА и в итоге образуется б-ти углеродная молекула (Зметилр-гидроксиглутарилКоА, фермент - р-гндрокир метлглюторилКоА-синтаза (ГМГ-синтетаза) Следующая реакция наиболее важная реакция этого синтеза, на которую направлены сегодня все ингибиторы синтеза холистерола (фермент является ключевым ферментом синтеза холистерола-ГМГ-редуктаза) Происходит восстановление до спиртовой группы и образуется соединение которое носит название мевалоновая кислота (монокарбоновая)
На втором этапе мевалоновая кислота в результате ряда последовательных превращений преобраз>ется в изопреноидные группировки. на 3 этапе из активных изопреноидных единиц путем последовательных реакций конденсаций образ> ется сквален, имеющий в своем составе 30 атомов углерода (т е используется по крайней мере 6 изопреноидных группировок) На 4 этапе идет циклизация сквалена в соединение стероидной природы - ланоетерин, имеющий в своем составе, так же 30 углеродных атомов. Следует отметить, что некоторые промежуточные продукты этого синтеза используются для синтеза других соединений, в частности коэнзимаО (источник энергии д/ja переноса электронов и протонов т е •это компонент главной дыхательной цепи митохондрий) дошхомЬос&ат (принимает участие в синтезе гетероо'шгосахаридных компонентов в составе гликопротеидов) Ключевая рюль в регуляции синтеза холистерола в клетках принадлежит ферменту ГМГ-КоАредуктазе
При повышении содержания хояистерола в клетках, В независимости от того синтезирован он здесь в клетках или поступил из вне происходит снижение активности этого фермента, причем установлено что в данном случае речь идет не о прямом влиянии холистерола на активность фермента, а в основе ингибирующего действия лежат другие механизмы.
Ацетил КоА
В кишечной стенки всосавшиеся ацилгицерины распадаются под действием тканевых липаз с образованием свободных жирных кислот и глицерола Часть моноацилглицеринов может превращаться в триащгаглицерины без предварительного расшепления йо так называемый моноацклглицериновый путь ресинтеза Все высшие жирные кислоты всосавшиеся к, кишечника используются в энтероцитах для ресинтеза различных лилидов Но перед тем как: вклю иться в различные липиды высшие жирные кислоты должны быть активированы Процесс активации высших жирных кислот состоит из 2 этапов
1 этап. За счет взаимодействия жирных кислот и использования специального фермента образуется ацшюденилаты
(так называемый термодинамический контроль направления процесса)
ЖК + АТФ -> R-С-АМФ + пирофосфат расщ. до ФК.(термодинамический контроль)
2 этап Происходит образование активной жирной кислоты соединенной с КоА и высвобождение АМФ Образование ацилКоА катализируется специальным ферментом, причем он катализирует и первую и вторую реакцию ацилКо Чсиптетата (тиокиназа)
R-С-АМФ + НSКоА -> R-С-SКоА + АМФ
В ходе активации высшей жирной кислоты АТФ распадается до АМФ и 2 остатков фосфорной кислоты таким жирные кислоты участвуют в активированной форме.
19. Кетоновые тела.
Соединения ацетоуксусные и р-гидроксимасляные кислоты поступают в кровь, а затем идут в клетки тканей, но для этих молекул диффузионного барьера не существует, поэтому они служат эффективным энергетическим топливом. Эти соединения получили название - ацетоновые тела. К ацетоновым телам относится и сам ацетон (диметилкетон) В то же время в гепатоциты высшие жирные кислоты поступают минуя диффузионный барьер потому, что гпатоциты в печеночных синусах непосредственно контактируют с кровью.
Биосинтез и распад ацетоновых тел. Жирные кислоты поступающие в гепатоциты, активируются и подвергаются р-окислению с образованием ацетилКоА Именно этот ацетилКоА используется для синтеза ацетоновых тел, согласно схеме В ходе первой реакции (в первую реакцию вступают 2 молекулы ацетилКоА, фермент ацетилКоА-ацетилтрансфераза = тиолаза) образуется 4-х углеродная молекула ацетоацетилКоА Эти соединения макроэргические поэтому в этом синтезе не принимает участие АТФ Входе следующей реакции (фермент В-гидрокси-ВметилглюкоилКоА-синтетаза- первые этапы биосинтеза ацетоновых тел и холестерина абсолютно равнозначны Это одна из ключевых реакций синтеза ацетоновых тел) используется еще одна молекула ацетилКоА, вода Образуется б-и углеродная молекула - р-гидроксир-метилглютарилКоА. Последняя реакция - лиазная (катализирует фермент ГМГ-лиаза), происходит отщепление ацетилКоА и образование 4-х углеродной молекулы - ацетоацетата.
Как образуются два других соединения, относящихся к группе ацетоновых тел? Из ацетоуксусной кислоты спонтанно, чаще всего, или иногда за счет декарбоксилазы происходит отщепление карбоксильной группы в виде углекислого газа и образуется ацетон Ацетоуксусная кислота восстанавливается в ходе реакции катализируемой ферментом р-гидроксибутератдегидрогиназой с использованием НАД+Н+, в итоге образуется р-гидроксимасляная кислота Это третий составной элемент ацетоновых тел Образовавшиеся ацетоновые тела поступают из гепатоцитов в кровь и разносятся к клеткам Процесс синтеза ацетоновых тел идет постоянно и ацетоновые тела всегда присутствуют в крови в концентрации 30мг/л. При голодании их содержание может увеличиваться до 400-500 мг/л Еще больше концентрация при сахарном диабете в тяжелой форме до 3000-4000 мг/л
Ацетоновые тела в норме хорошо утилизируются клетками периферических тканей, в особенности это касается скелетных мышц и миокашш Ске,иетные мыишы и миокаод значительную часть нужной им энергии получают за счет окисления ацетоновых тел Только нервные клетки в обычных условиях не утилизируют ацетоновые тела, однако при голодании даже головной мозг 50-75% соей потребности в энергии удовлетворяет за счет окисления ацетоновых тел. Ацетоацетат, поступающий в клетки различных тканей, прежде всего подвергается активации помощью одного из двух механизмов Ацетоацетат с участием фермента тиокиназы, за счет энергии АТФ превращается в ацетоацетилКоА
Второй путь, является превалирующим в активации, это за счет фермента тиофоразы Реакция, в которой принимают участие сукценнКоА и адетоацетат, приводит к образованию ацетоацетилКоА и образование сукцината, Образующийся ацетоацетилКоА далее дает 2 молекулы ацетилКоА (принимает участие HSKoA, это тиолазная реакция) АцетилКоА поступает в цикл Кребса, где ацетильные остатки окисляются до углекислого газа и воды Ацетоновые тела по значимости - 3 тип топливной энергии В гепатоцитах нет фермента тиофоразы, поэтому образовавшийся в гепатоцитах ацетоацетат не активируется и не окисляется Таким образом печень экспортирует ацетоацетат, другими словами синтезирует этот вид топлива для других клеток р-гидрокснбутерат окисляется путем дегидрироания в ацетоацетат, дальше ацетоацетат в ацетилКоА Что касается ацетона, возможно 2 варианта окисления Дело в том, что ацетон очень летуч поэтому большое количество выделяется вместе с выдыхаемым воздухом, кроме того ацетон выделяется с водой 1 путь Ацетон расщепляется до ацетильного и формильного остатка 2 путь Через пропандиол он превращается в пируват
Ацетоновые тела накапливаясь в крови и тканях оказывают ннгибирующие действие на липолиз, в особенности это касается расщепление триглицеридов в липоцитах Дело в том, что избыточное накопление в крови ацетоновых тел приводит к развитию ацидоза Снижение уровня липолиза в клетках жировой ткани приводит к уменьшению притока жирных кислот в гепатоцига, к снижению скорости образования ацетоновых тел н следовательно к снижению содержания в крови.
Транспортные формы лнпидов
Все липиды присутствующие в крови входят в составе смешанных надмолекулярных белок-липидных
комплексов. причем ВЖК связаны с альбуминами крови, все другие липиды входят в состав липопротеидов плазмы крови С месь всосавшихся и ресентезированиых в кишечнике липидов поступает в лимфатическую систему а затем через грудной лимфатический проток в кровь и распространяется током крови по клеткам и тканям Поступление липидов в лимфу наблюдается уже через 2 часа после приема пищи Элементарная гиперлипидемия ^повышение концентрации яипидов в крови) достигает максимума через б 8 часов после приема пищи, а через 10 12 часов после приема она полностью исчезает Как же это все происходит? Трнглицериды фосфолипиды и холестирол
Перенос этих соединении осуществляется особым образом организованных надмолекулярных агрегатов получивших название -липопротеидные частицы (ЛП). В состав липопротеидов могут входить молекулы липидов различных классов и молекулы белков Все ЛП имеют общий план структуры
Во внешней оболочке или так называемый внешний мономолекулярный слой липопротеидные частицы образуют белки (их называют апобелки или апопротеины) свободный холистерол и фосфолипиды Причем гидрофильные участки этих молекул обращены кнаружи и контактируют с водой, гидрофобные участки располагаются кнутри т е в сторону ядра Ядра гидрофобных липопротеидньгх частиц образуют прежде всего триглицериды далее этерефицированный холистерол, кроме того сюда могут включаться жирорастворимые витамины или другие гидрофобные молекулы Их подразделяют на 1) Хиломикрокы (ХМ) 2) Липопротеиды очень низкой плотности 3) Лкпопротеиды низкой плотности 4) Липопротеиды высокой плотности
В транспорте экзогенных липидов те липндоБ поступающих во внутреннюю среду организма из кишечника принимают главным образом два вида липопротеидов ХМ и Липопротеиды очень низкой плотности (лонп) Содержание липидов в ряду от ХМ до ЛПВП снижается, содержание белков нарастает постепенно возрастает содержание фосфолипидов возрастает и только содержание холистерола до липопротеидов низкой плотности увеличивается, но затем при переходе в лпвп оно снижается. Ведущую роль в транспорте экзогенных тлипидов играют хнпомикроны.
Матболизм ХМ.
Липопротеид липаза расщепляет триглицериды хиломикронов до глицерола и высших жирных кислот. Часть высших жирных кислот поступает в клетки а другая часть связывается с альбуминами и уносится током крови в другие ткани. Глицерин так же может утилизироваться либо в клетках непосредственно данного органа либо уносится током крови. Кроме триглицеринов хиломикронов ЛП липаза расщепляет также триглицериды липидов очень низкой плотности ХМ после атаки липопротеидлигаз потеряв значительную часть липидов превращаются в ремнантные хиломикроны (остаточные ХМ они по размерам меньше) Эти ремнантные ХМ захватываются рецепторами печени где они полностью расщепляются, а часть ХМ превращается путем сложных перестроек в липопротеиды высокой плотности В норме спустя 10-12 часов после приема пищи плазма практически не содержит ХМ.
Метаболизм ЛПВП.
Общий пул лпвп в циркулирующей крови формируется за счет трех источников 1 Синтез лпвп в печени 2 Образование лпвп нз ремнантных ХМ 3 Синтез в тонком кишечнике 4 Преимущественно теп синтезируются в печени Наиболее популярной точкой зрения в отношении биологической роли лпвп является следующая теп
этот холистерол в печень или кишечник. В печени этот холестерол: превращается в желчные кислоты 2 часть секретируется с желчью Поступившый в стенку кишечника холистерол с лпвп используется для синтеза 1) ХМ 2 лпонп и в то же время 3 часть холистерола может секретироваться в просвет кишечника.
В любом случае чтановится ясно, чтофункционирование лпвп будет способствовать выведению лишнего холестерола
из кишечника. Лпвп - антиатерогенные липопротеидами, в отличии рассмотренные ранее лпонп и лпнп атерогенные липопротеиды (высокая концентрация которых несомненно стимулируют формирование атеросклероза и
соотвествующих осложнений.
Спиртовое брожение
Реакция спиртового брожения подобна гликолизу. Расхождение начинается только после образования пирувата.Конечный этап гликолиза заменяется двумя ферментативными реакциями. Сначала пируват подвергается декарбоксилированию, продуктом которого является ацетальдегид. Данная реакция происходит при участии пируватдекарбоксилазы, ТПФ и ионов магния. 
После ацетальдегид восстанавливается водородом, который отщепляется от кофермента НАДН. При этом ацетальдегид восстанавливается до этанола. Собственно, цель спиртового брожения — это окисление NADH, чтобы он мог снова принять участие в гликолизе. Катализатором является алкогольдегидрогеназа. 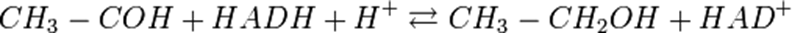
Таким образом, продуктами спиртового брожения являются этанол и, а не молочная кислота, как в молочнокислом брожении.
В результате получается реакция: 
26.Аэробный распад глюкозы. Схема. Пути регенерации НАД+. Энергетический эффект. Регуляция аэробного распада глюкозы. Эффект Пастера.
Аэробный распад глюкозы можно выразить суммарным уравнением:
С6Н12О6 + 6О2 → 6СО2 + Н2О + 2880 кДж/моль.
Этот процесс включает несколько стадий:
1. Аэробный гликолиз - процесс окисления глюкозы с образованием двух молекул
пирувата;
2. Общий путь катаболизма, включающий превращение пирувата в ацетил-КоА и его
дальнейшее окисление в цитратом цикле;
3. Цепь переноса электронов на кислород, сопряжённая с реакциями дегидрирования,
происходящими в процессе распада глюкозы.

Эффект Пастера.
Снижение скорости потребления глюкозы и прекращение накопления лак-тата в присутствии кислорода носит название эффекта Пастера. Впервые это явление наблюдал Л. Пастер во время своих широко известных исследований роли брожения в производстве вина. В дальнейшем было показано, что эффект Пастера наблюдается также в животных и растительных тканях, где кислород тормозит анаэробный гликолиз. Значение эффекта Пастера, т.е. перехода в присутствии кислорода от анаэробного гликолиза или брожения к дыханию, состоит в переключении клетки на наиболее эффективный и экономичный путь получения энергии. В результате скорость потребления субстрата, например глюкозы, в присутствии кислорода снижается. Молекулярный механизм эффекта Пастера заключается, по-видимому, в конкуренции между системами дыхания и гликолиза (брожения) за АДФ, используемый для образования АТФ. Как известно, в аэробных условиях значительно эффективнее, чем в анаэробных, происходят удаление Piи АДФ, генерация АТФ, а также регенерирование НАД+, окисленного из восстановленного НАДН. Иными словами, уменьшение в присутствии кислорода количества Рi и АДФ и соответствующее увеличение количества АТФ ведут к подавлению анаэробного гликолиза.
Первая реакция.
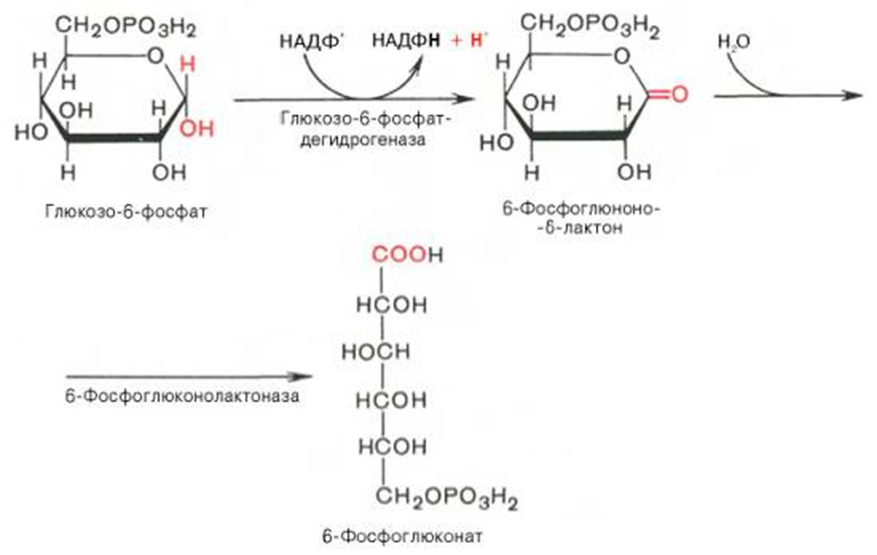
Вторая реакция.


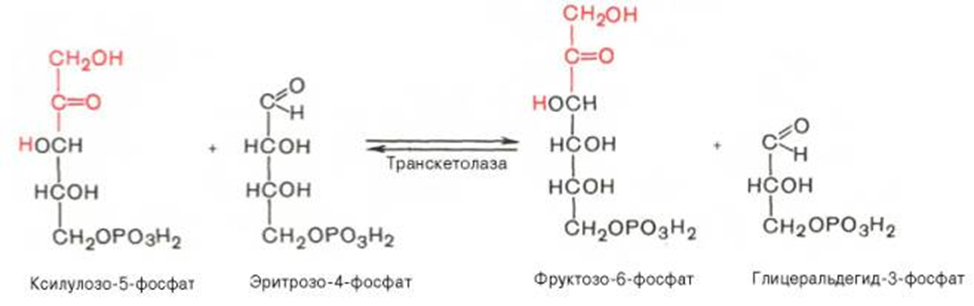
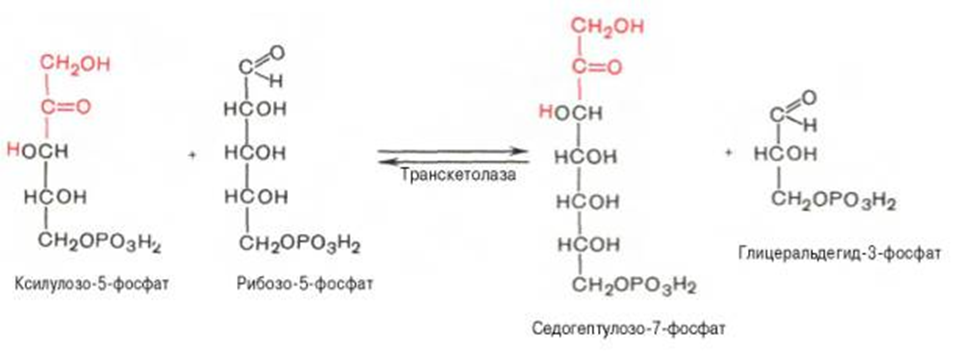
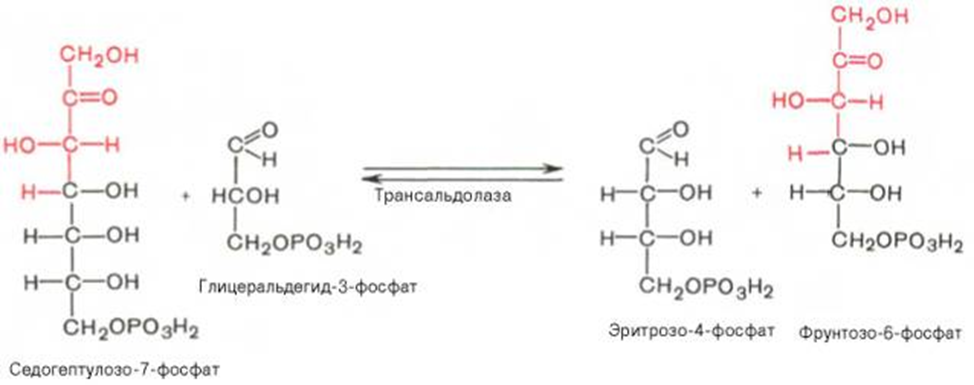
Биороль: Значение этого пути в обмене веществ велико. Он поставляет восстановленный НАДФН, необходимый для биосинтеза жирных кислот, хол






