Стимулом для исследования реакции микроорганизмов на облучение послужило стремление решить проблемы, связанные с опасностью радиации для человечества. Хорошо известно, что излучения разных типов обладают потенциальной способностью оказывать на живые организмы разрушительное воздействие.
Однако, если не говорить о высоких дозах, излучения во внешней среде носят такой характер, что для любой клетки существует определенная вероятность избежать повреждения. Исходя из этого, можно было бы предположить, что одноклеточным организмам удается выйти из опасного положения благодаря тому, что они очень быстро размножаются. Тем не менее это, по-видимому, не так, поскольку у них выработались дополнительные средства защиты от летального или повреждающего воздействия облучения. Одноклеточные организмы располагают множеством защитных механизмов, причем многие виды используют не один, а большее число способов борьбы с радиационными повреждениями.
Клеточные механизмы защиты от повреждающего действия радиации настолько широко распространены у микроорганизмов, что трудно оценить опасность, которую представляет для них этот фактор среды. Только тогда, когда защитные механизмы почему-либо не действуют, становятся очевидными реальные размеры опасности, исходящей в основном от коротковолновой части солнечного спектра. Не удивительно поэтому, что начало выяснению природы радиорезистентности было в значительной мере положено обнаружением радиационно-чувствительных мутантов. Такие мутанты не только позволили выявить опасность радиации, но и послужили средствами для исследования механизмов, сдерживающих эту опасность.
Инактивация клеток, вызываемая облучением в относительно низких дозах, обусловлена главным образом повреждением ДНК. Это заключение подтверждается тем, что у прокариот большинство изученных радиационно-чувствительных мутантов дефектны по функциям, имеющим отношение к ДНК. Кроме того, исследование таких мутантов показало, что резистентность обычно достигается не в результате защиты ДНК от индукции повреждении, а скорее благодаря действию механизмов, репарирующих ДНК после того, как повреждения возникли.
Адаптация микроорганизмов к высоким дозам излучений основана на механизмах, участвующих в исправлении повреждений, вызываемых облучением. Известны три независимые системы репарации повреждений ДНК, индуцируемых облучением. Одна из них представляет собой обратную фотохимическую реакцию, происходящую под действием видимого света и фотореактивирующего фермента; вторая — вырезание и замещение поврежденного участка ДНК до ее репликации, а третья — пострепликативную репарацию. Первый из упомянутых механизмов действует только на пиримидиновые димеры, индуцируемые ионизирующим излучением. Многие организмы для защиты от неблагоприятного воздействия радиации используют все три системы. Несмотря на то, что репаративные механизмы имеются у всех исследованных в этом отношении микроорганизмов, последние различаются по своей способности переносить облучение. Самый яркий пример такой вариабельности — исключительно высокая резистентность М. radiodurans и родственных ему видов.
Парадоксально, но одно из последствий облучения — возникновение мутаций — может дать организму преимущество при отборе. Поэтому есть основания предполагать, что с эволюционной точки зрения для организма выгодно установление некоторого равновесия между резистентностью и чувствительностью к радиации; возможно, именно по этой причине защита никогда или почти никогда не бывает полной. Следует ожидать, что соотношение между чувствительностью и резистентностью к облучению неодинаково у разных организмов; и действительно, среди различных видов наблюдается исключительное разнообразие по степени их резистентности к летальному и мутагенному действию облучения. Такое разнообразие создает большие возможности для исследования явлений резистентности и чувствительности организмов к радиации.
Клеточные механизмы, обеспечивающие радиорезистентность, можно разделить на две большие группы. К первой относятся системы, предотвращающие возникновение повреждений в клетке. Вторая включает механизмы, которые восстанавливают (репарируют) повреждения в ДНК, индуцируемые облучением. В настоящем обзоре будут рассмотрены оба аспекта радиорезистентности, но второму из них — репарации — в настоящее время уделяется значительно больше внимания и соответственно он будет обсуждаться здесь более детально. Исследования в этой области были стимулированы одним удивительным открытием: выяснилось, что по крайней мере некоторые из путей репарации более или менее независимы от основных процессов клеточного метаболизма. Поэтому оказалось возможным выделить дефектные штаммы, у которых нарушена система репарации индуцируемых облучением повреждений, но которые тем не менее остаются жизнеспособными. Такие мутанты чувствительны к облучению и в качестве генетических инструментов оказываются исключительно полезными в исследовании клеточных механизмов репарации. Работы в этой области существенно углубили понимание природы радиорезистентности.
Диапазон доз облучения, которому микроорганизмы могут подвергаться эпизодически или постоянно, значительно расширился в последние годы в связи с созданием искусственных источников радиации. В результате этого микроорганизмы стали испытывать воздействие повышенного уровня радиации в окружающей среде. Реакция на такое воздействие представляет как практический, так и научный интерес. Например, при использовании высоких доз облучения для стерилизации пищи возникает проблема, связанная с возможностью индукции или селекции радиорезистентных микроорганизмов, что может иметь неблагоприятные последствия.
Излучение в окружающей среде подразделяется на ионизирующее и неионизирующее. Оба вида опасны для микроорганизмов, но из всех естественных излучений неионизирующая солнечная радиация обладает наибольшим потенциалом биологически вредного воздействия.
Одним из наиболее резистентных к ультрафиолетовому излучению микроорганизмов является Bodo marina, относящийся к морским жгутиковым.
Для инактивации 90% клеток этого организма требуется доза 112 000 эрг-мм-2. Отмечено, что простейшие вообще более резистентны к облучению, чем бактерии (дозы, инактивирующие 90% клеток, колеблются от 5000 до 12 000 эрг-мм-2 для простейших и от 4 до 250 эрг-мм-2 для бактерий). Резистентность разных видов бактерий варьирует в очень широких пределах (Камшилов М.М., 1974).
В отношении резистентности организмов к ионизирующему излучению наблюдается сходная картина. Так, дозы, убивающие 50% клеток в культурах Е. coli, дрожжей, амебы, В. mesentericus и инфузорий возрастают от 5600 до 350 000 Р.
Имеется корреляция между резистентностью организма и уровнем радиации в среде его обитания. Так, микроорганизмы, выделенные из радиоактивных минеральных источников, в 3— 10 раз более резистентны к радиации, чем организмы тех же видов, выделенные из нерадиоактивной воды.
Обнаружен вид Pseadomonas, обитающий в ядерных реакторах, где средняя доза излучения, по-видимому, превышает 106 ФЭР (физический эквивалент рентгена).
В связи с такими случаями явной адаптации микроорганизмов к радиоактивному излучению был поднят вопрос о том, насколько вероятно появление радиорезистентных организмов при использовании облучения для стерилизации. Действительно, Micrococcus radiodurans, обладающий наиболее высокой радиорезистентностью из всех изученных бактерий, был первоначально обнаружен в консервированном мясе, которое подвергалось Y-облучению в дозе несколько мрад. Этот организм может переносить дозы облучения до 500 крад без какой-либо заметной инактивации (Кашнер Д., 1981).
Более устойчивы к излучению грамположительные микробы и менее устойчивы грамотрицательные. Повышенная устойчивость к излучениям отмечена у клостридий ботулизма: они погибают только после воздействия на них дозами в 25—40 кГр. Для достижения стерильности в некоторых случаях необходимо излучение в 50 кГр. Устойчивы к излучению и риккетсии; их устойчивость примерно такая же, как и у спор бацилл.
Некоторые микробы (возбудитель сибирской язвы, кишечная палочка и др.) приобретают устойчивость к излучениям. После нескольких облучений она у них повышается в два или более раза (Асонов Н.Р., 1997).
Защитные механизмы
Одним из универсальных механизмов адаптации к световому излучению высокой интенсивности и защиты от токсичных форм фотосенсибилизированного кислорода является синтез каротиноидных пигментов. Характерным примером может служить яркая окраска микроорганизмов, живущих в условиях высокой освещенности (в воздухе, на поверхности скал, обнажений горных пород, в высокогорье и т.д.) (Заварзин Г.А., 2001).
Ранние исследования радиационной резистентности были направлены в первую очередь на поиски внутриклеточных веществ, защищающих организм от повреждений. В настоящее время внимание исследователей концентрируется в основном на механизмах, тем или иным способом исправляющих повреждения в ДНК, индуцируемые облучением. Эти механизмы имеют важное значение. Тем не менее представляется вероятным, что определенную вспомогательную роль играют и защитные механизмы.
Для радиорезистентных организмов обычно характерна усиленная пигментация, что чаще всего является причиной резистентности. Пигменты действуют как «энергетические ловушки», препятствующие радиации или ее продуктам достигать ДНК или любых других жизненно важных мишеней.
Резистентность может быть обусловлена также присутствием определенных продуктов метаболизма (внутриклеточных радиопротекторов). Так, Е. coli более резистентна при облучении в присутствии экстрактов из М. radiodurans, чем при облучении в буферном растворе. Защитное действие экстрактов из М. r аdiodu r ans связано с уменьшением выхода радикалов при участии механизма, аналогичного тому, который действует в отношении известных химических веществ-протекторов.
Своеобразный метаболизм серосодержащих аминокислот у М. radiodurans позволяет думать, что эти аминокислоты выполняют роль сульфгидрильных веществ-протекторов. Зависимость радиорезистентности от концентрации экстракта имеет двухкомпонентный характер: в низких концентрациях он оказывает на тест-организм сенсибилизирующее действие, а в высоких концентрациях — защитное.
Радиорезистентность может определяться уровнем каталазной активности в. клетке. Было показано, что для некоторых бактерий с повышенной радиорезистентностью характерно высокое содержание каталазы.
С возрастанием радиорезистентности увеличивается длина клеток: значительная часть клеток наиболее резистентных штаммов была в 30—40 раз длиннее нормальных. У клеток этих штаммов наблюдалось также своеобразное явление почкования. У резистентных штаммов, упомянутых выше, удлинение клеток было устойчивым признаком, наблюдавшимся в течение трех лет.
К важным факторам, от которых зависит реакция той или иной клеточной системы на любой физический или химический агент, относится состав клеточной стенки. В случае химических мутагенов структура клеточной стенки может определять ее проницаемость, влияя, таким образом, на чувствительность клетки к данному агенту. Хотя структура клеточной стенки не оказывает влияния на проникающую способность ионизирующего излучения, она тем не менее может иметь значение для радиорезистентности микроба. Например, вполне вероятно, что какой-либо связанный с мембраной ферментный комплекс, освобождающийся или активируемый под действием радиации, играет определенную роль в системе (системах) репарации или обусловливает конечную инактивацию клетки. Ионизирующее излучение вызывает освобождение связанной с клеточной поверхностью экзонуклеазы у М. radiodurans. При облучении в дозе 400 крад, сублетальной для этого организма, в клетках остается только 10% фермента, причем степень освобождения фермента зависит от дозы облучения.
Микробы-сапрофиты более устойчивы у световому излучению в сравнении с патогенными. Это объясняется тем, что они чаще подвергаются действию прямых солнечных лучей, поэтому являются более адаптированными (Радчук Н.А., Дунаев Г.В., 1991).
Увеличение содержания ДНК в клетке служит одним из факторов ее радиорезистентности. Это может быть обусловлено либо увеличением числа нуклеоидов в клетке, либо ее полиплоидностью. Нитевидная форма резистентных клеток Е. coli позволяет предполагать, что в них реализуется первый механизм. Но содержание ДНК в клетках этих штаммов практически не отличается от такового в клетках дикого типа.
Определение содержания GC-nap в ДНК восьми видов бактерий показало, что существует обратная зависимость между GC-содержанием и резистентностью клеток к рентгеновским лучам. В то же время между GC-содержанием и резистентностью к УФ-облучению наблюдается прямая зависимость. Такая корреляция утрачивает какой бы то ни было смысл в случае М. radiodurans, резистентного к обоим типам излучения; однако она может иметь некоторое значение при отсутствии у бактерий эффективных систем репарации. Действительно, ДНК М. radiodurans характеризуется тем же нуклеотидным составом, что и ДНК штаммов Pseudomonas, исключительно чувствительных к ионизирующей радиации (Покровский В.И., 1999).
Механизмы репарации ДНК
В основе радиорезистентности бактерий лежат разнообразные внутриклеточные процессы, участвующие в репарации поврежденной ДНК. Большую ценность для исследования этих процессов представляет наличие хорошо охарактеризованных мутантных штаммов, радиационная чувствительность которых варьирует в чрезвычайно широких пределах.
При помощи генетических скрещиваний были получены двойные и тройные мутанты дрожжей, у которых репаративная активность полностью отсутствует. Сравнительное исследование штамма дикого типа и сверхчувствительных двойных и тройных мутантов S. сеrevisiae показало, что если нормальный штамм довольно легко переносит образование в ДНК почти 16 000 димеров (37% выживания), то двойные и тройные мутанты остаются резистентными в присутствии не более 50 и 1 димера соответственно. Пониженная резистентность таких двойных и тройных мутантов служит убедительным свидетельством в пользу существования различных путей репарации радиационных повреждений.
В зависимости от того, участвует ли видимый свет в модификации повреждений ДНК, репарацию можно подразделить на световую и темновую. Конкретно под световой репарацией понимается феномен фотореактивации, впервые описанный у актиномицетов. Механизм фотореактивации действует только на пиримидиновые димеры. В этом процессе участвует фермент фотореактивации, который связывается с димерами. Образующийся фермент-субстратный комплекс активируется видимым светом, что приводит к мономеризации димеров in situ. Таким образом, летальный эффект УФ-облучения существенно снижается, если облученные клетки подвергаются затем воздействию видимого света с длинами волн от 360 до 420 нм (см. рис. 6.1).

Рис. 6.1 Световая репарация ДНК
Фотореактивация служит мощным инструментом исследования летальных и мутационных повреждений, так как их репарация под влиянием света может быть использована в качестве критерия для решения вопроса о том, обусловлена ли инактивация ДНК образованием пиримидиновых димеров.
К другому типу реактивации клеток видимым светом относится его защитное действие. В этом случае увеличение выживаемости клеток наблюдается при освещении их видимым светом перед УФ-облучением. Этот феномен объясняют тем, что видимый свет индуцирует задержку клеточного деления. В результате такой задержки остается больше времени для репарации повреждений, вызываемых УФ-облучением (см. рис. 6.2).
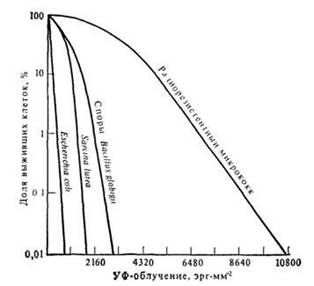
Рис. 6.2 Зависимость выживания клеток бактерий от величины облучения
Под «темновой репарацией» понимают репарацию без участия света. В настоящее время известны две системы такого типа: эксцизионная репарация и пострепликативная рекомбинационная репарация. Репарация первого типа требует присутствия ферментов, которые узнают нарушения структуры ДНК, удаляют затронутые участки, замещая их нормальными нуклеотидными последовательностями, и, наконец, восстанавливают первоначальную структуру ДНК, замыкая полинуклеотидную цепь (см. рис. 6.3).

Рис. 6.3Темновая репарация ДНК
Действие разнообразных инактивирующих агентов на клетки может приводить к возникновению в ДНК целого ряда различных повреждений. Детальное изучение системы эксцизионной репарации стало возможным благодаря наличию радиационно-чувствительных мутантов, с помощью которых удалось выделить и охарактеризовать специфические ферменты, принимающие участие в разных стадиях этого процесса. У Е. coli имеется, по крайней мере, четыре таких этапа. На первом этапе происходит разрыв цепи ДНК вблизи повреждения под действием эндонуклеазы, узнающей нарушения структуры ДНК. Такая УФ-специфическая эндонуклеаза была выделена из Micrococcus luteus и Е. coli. За разрывом цепи ДНК следует удаление пиримидиновых димеров, осуществляемое экзонуклеазой. Удаление димеров сопровождается дополнительной деградацией ДНК с образованием брешей, размеры которых варьируют от 20 до 400 нуклеотидов. Затем бреши заполняются с помощью ДНК-полимеразы, использующей в качестве матрицы интактную комплементарную цепь ДНК. Заключительный шаг в этой последовательности событии состоит в восстановлении целостности полинуклеотидной цепи в результате сшивания разрыва лигазой.
Второй тип темновой репарации — пострепликативная рекомбинационная репарация — был впервые описан Говард-Флендерсом. Как указывает само название, эта репаративная система устраняет повреждения в ДНК после того, как произошла ее репликация.
Клеточная система репликации способна «обходить» некоторые из димеров в матричной цепи ДНК, оставляя в растущей цепи бреши, расположенные напротив каждого из них. Число брешей, возникающих таким путем, примерно соответствует числу димеров в ДНК. В результате процесса, сходного с рекомбинацией и включающего обмен между сестринскими нитями, образуется ДНК с двумя интактными цепями. Обнаружено, что при заполнении брешей происходит обмен между облученной родительской цепью ДНК и необлученной дочерней цепью. Установлено, что для образования интактных, не содержащих димеры молекул ДНК, заполнение брешей не обязательно, вместо этого концентрация димеров может просто постепенно снижаться в ходе последовательных циклов репликации ДНК после облучения (Покровский В.И., 1999).






