

Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Топ:
Техника безопасности при работе на пароконвектомате: К обслуживанию пароконвектомата допускаются лица, прошедшие технический минимум по эксплуатации оборудования...
Марксистская теория происхождения государства: По мнению Маркса и Энгельса, в основе развития общества, происходящих в нем изменений лежит...
Теоретическая значимость работы: Описание теоретической значимости (ценности) результатов исследования должно присутствовать во введении...
Интересное:
Национальное богатство страны и его составляющие: для оценки элементов национального богатства используются...
Влияние предпринимательской среды на эффективное функционирование предприятия: Предпринимательская среда – это совокупность внешних и внутренних факторов, оказывающих влияние на функционирование фирмы...
Распространение рака на другие отдаленные от желудка органы: Характерных симптомов рака желудка не существует. Выраженные симптомы появляются, когда опухоль...
Дисциплины:
|
из
5.00
|
Заказать работу |
Содержание книги
Поиск на нашем сайте
|
|
|
|
Существует возможность количественно оценить клады различных АО в МО. Как уже отмечалось, используя разложение МО по линейным комбинациям АО ji= åсimcm(2.12), удобно ввести матрицу плотности (матрицу зарядов-порядков связей) P c элементами Рmn = åj сjmсjn(суммирование ведется по занятым МО). Энергия молекулы с закрытыми оболочками записывается с помощью этой матрицы весьма компактно:
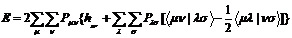 (3.5)
(3.5)
Матрица плотности описывает распределение электронной плотности по молекуле следующим образом (закрытые оболочки):
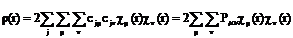 (3.6)
(3.6)
Перепишем последнее выражение таким образом, чтобы выделить вклады АО атомов I и J,
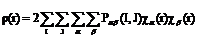 ,
,  aÎI,bÎJ (3.7)
aÎI,bÎJ (3.7)
и проинтегрирум левую и правую части (3.7). Учитывая, что АО нормированы, имеем:
 (3.8)
(3.8)
(интеграл перекрывания Sab(I,J)=1, если a=b, I=J). Таким образом, полное число электронов молекулы Nв приближении МО ЛКАО оказывается формально распределенным по атомам и связям между ними. Величина Рaa(I) есть электронная заселенность орбитали a, центрированной на атоме I, а 2Рab(I,J)Sab(I,J) –электронная заселенность перекрывания орбиталей aи b, центрированных на атомах Iи J.
Малликен предложил оценивать электронные заселенности атомов, деля заселенности перекрывания орбиталей между рассматриваемой парой атомов поровну.
Тогда полное число электронов, приписывемое атому I, равно
 , I<J, a<b (3.9)
, I<J, a<b (3.9)
Полный заряд на атоме qI(эффективный заряд) вычисляется как разность
qI= ZI- QI, (3.10)
где ZI– заряд ядра атома I.
Ясно, что описанный метод анализа электронных заселенностей орбиталей дает лишь приближенную оценку распределения заряда по молекуле: получаемые величины зависят от используемых при расчете метода и базиса, а деление электронов между атомами без учета их природы неправомочно. Кроме того, в ортогональном базисе все электроны оказываются формально распределенными только между атомами. Тем не менее, анализ заселенностей орбиталей по Малликену проводится при выдаче результатов расчета по всем современным квантовохимическим программам.
Величины 2Рab, следуя Коулсону, называют орбитальными зарядами, а 2Рab-
порядками связей: отсюда и происходит название матрицы Р. Знак 2Рabопределяет конструктивная или деструктивная интерференция имеет место при взаимодействии данной пары АО. Полный порядок связи между атомами находят как сумму вкладов от перекрывания соответствующих АО.
Используя матрицу зарядов-порядков связей, легко вычислить дипольный момент молекулы:
 (3.11)
(3.11)
Дипольный момент нейтральной молекулы не зависит от выбора начала отсчета.
Дипольные моменты молекул определяются измерением комплекной диэлектрической проницаемости как функции частоты и температуры, из ИК спектров и с помощью других методов. Сравнивая результаты расчета и измерений, можно оценить надежность проведенного исследования.
Макромолекулы как основа организации биологических структур. Основу структурной организации живого составляют макромолекулы, прежде всего важнейшие биополимеры – белки и нуклеиновые кислоты. В макромолекулах происходят сложные процессы трансформации Е, включающие миграцию Е электронного возбуждения и транспорт электронов. В основе функционирования макромолекул лежит электронно-конформационные взаимодействия, которые проявляются в самых различных процессах, где участвуют биологически активные макромолекулярные комплексы. Тепловые движения атомов, входящие в полипептидную цепь, их повороты и вращения вокруг связей приводят к созданию большого количества внутренних степеней свободы. Это приводит к свертыванию цепи и образованию клубков макромолекул – макроскопические системы, в которых проявляются статический характер параметров (размеры, формы, степень свертывания). Как физический объект их своеобразие проявляются в сочетании как статических, так и механических особенностей поведения макромолекул. С одной стороны большое число взаимодействий атомов создают большое количество степеней свободы и возможны создания различных конформаций, с другой стороны наличие химической связи между атомами ограничено -возможно образование конформеров. Взаимодействия атомов химической ковалентной связи определяет: 1. цепное строение биополимеров. 2. соединение друг с другом мономеров. Клетки и их органоиды – гетерогенные системы. Их существование и функц-ие опред-ся межмолекулярными взаимодействиями нековалентного характера (взаимодействуют слабо, потому что сильные взаимодействия создали бы устойчивые жестские структуры, лишенные молекулярной подвижности, а молекулярная подвижность необходима для выполнения различных задач (регуляция химических реакций, трансформация Е). Слабое взаимодействие в биологических системах: 1.Вандервальсовы силы, 2.Ионные связи, 3.Водородные связи, 4.Гидрофобные взаимодействия. Первичная структура – линейная. Важную роль в конформации полипептидов играют вандевальсовые силы, гидрофбные взаимодействия, водородные связи. Вторичная структура. Пептидная цепь в белках имеет спиральную конфигурацию (-спираль). Каждый атом H2 имеет избыточный «+» заряд, притягивающийся к «-» заряженному атому О2 в следующем витке спирали. Внутри образуются пептидные связи, а боковые радикалы аминокислот обращены наружу и могут взаимод-ть с молекулами окружающей среды. Спиральная конфигурация – вторичная. Структура полипептидной цепи спирализована неполностью. Инсулин – 60%. Причиной нарушения спирали являются: 1. Образование дисульфидных связей, которые могут соединить несколько спиралей между собой. В местах образования их ослабляется Н-связь и нарушается спирализация. 2.Наличие радикалов некоторых аминокислот, которые не укладываются в спираль и образуют отдельные складки, скрещивания водородных связей. Такие параллельно расположенные участки - - структура, - конфигурация представляет собой складчатую структуру, которая включает параллельные цепи, связанные водородной связью. Исследования показали, если в полипептидной цепи есть остатки Глу, Ала, Лей- образуются ά – спираль, а если Мет, Вал, Изолей, то - структура. В зав-ти от хар-ра вторичной структуры белки делятся на три группы: 1.Белки с преобладанием - структуры (гемоглобин, миоглобин). 2.Белки, упакованы по типу - структуры. 3.Смешанная вторичная структура. Третичная структура – строго упорядоченная в пространстве укладка спирали и несколько участков цепи. Каждый белок имеет свою конфигурацию. Это связано с тем, что свободные карбоксильные, гидроксильные, аминные и другие группы боковых радикалов, взаимодействующих между собой с образованием амидных сложных эфирных связей. Водородные связи соединяют остатки двух соседних цепей и образуются дусульфидные мостики. Это делает структуру полужесткой. Четвертичная структура: Объединение двух и более субъединиц. Третичная структура приводит к созданию сложной активной белковой молекулы. Гемоглобин: 4 глобулы. В случае глобулы полипептидная цепь свернутая в клубок – третичная. Наличие нескольких сшивок S-S накладывает ограничение на конформацию. Глобула формирует слабые взаимодействия (гидрофобные→ взаимодействия цепных молекул друг с другом). Собранные в пачки рассматриваются и образуются фибриллярные структуры, которые функционируют вне раствора и формируют надмолекулярные белковые структуры, которые состоят из большего количества макромолекул с невалентными взаимодействиями- это клеточные мембраны, хромосомы, глобулы, нити в молекулах. Нуклеиновые кислоты. Уотсон Крик – структура ДНК, которая представляет собой правовинтовую спираль, из двух полинуклеиновых цепей, при этом одна цепь обвита вокруг другой. Таким образом, пары гетероциклических оснований нах-ся внутри. Обе цепи соединены между собой Н-связями, которые возникают между гетероциклическими основаниями.
Общий характер объемных взаимодействий и влияние внешнего поля на стабильность конформации биополимеров. Фазовые переходы. Кооперативные свойства макромолекул. Различные типы объемных взаимодействий в макромолекулах. Водородные связи, силы Ван-дер-Ваальса и стабильность вторичной и третичной структур. Поворотная изомерия и энергия внутреннего вращения. Конформация полипептидной цепи.
Объѐмные взаимодействия Все взаимодействия между атомами независимо от их конкретной физической природы при формировании различных макромолекулярных структур и переходов между ними можно разделить на два типа: взаимодействия ближнего порядка между атомами соседних звеньев и дальние взаимодействия или объемные эффекты между атомами, которые хотя и отстоят по цепи далеко друг от друга, но случайно сблизились в пространстве в результате изгибания цепи. В стабилизации молекулы биополимера принимают участие следующие объѐмные взаимодействия: 1. Водородные связи
2. Электростатические силы
3. Ван-дер-Ваальсовы взаимодействия
4. Гидрофобные взаимодействия
2.3. Объёмное взаимодействие.
Переходы глобула - клубок в биополимерах.
Геометрические размеры задаются с помощью среднеквадратичного расстояния биополимера - h2. Внутренняя пространственная структура задаётся пространственным распределением плотности звеньев. Вследствие объёмного взаимодействия, число звеньев в пространстве может меняться от точки к точке.
В полимерных нитях, вследствие взаимосвязанности звеньев, изменение плотности в одной точке пространства связано с изменением плотности в другой точке, то есть существует пространственная корреляция плотности. Если в макромолекуле отсутствует объёмное взаимодействие, то она не имеет достоверной пространственной структуры. В этом состоянии флуктуация (изменение вероятности) плотности имеет значение того же порядка, что и сама плотность. Такое состояние носит название клубка. Радиус корреляции 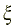 , то есть характерное расстояние, в пределах которого плотность звеньев резко меняется, становится того же порядка, что и размеры макромолекулы R:
, то есть характерное расстояние, в пределах которого плотность звеньев резко меняется, становится того же порядка, что и размеры макромолекулы R:
( ~R, R~ 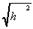 ,
, 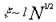 ).
).
Наличие объёмных взаимодействий может привести к такому состоянию, в котором флуктуация плотности мала по сравнению с самой плотностью. Такое плотное образование называется глобулой. В нем радиус корреляции флуктуации плотности намного меньше размеров молекулы <<R. Глобула в отличие от клубка обладает компактной пространственной структурой. Сердцевина большой глобулы примерно однородна, с постоянной концентрацией звеньев n0.
|
|
|

Двойное оплодотворение у цветковых растений: Оплодотворение - это процесс слияния мужской и женской половых клеток с образованием зиготы...

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...

Наброски и зарисовки растений, плодов, цветов: Освоить конструктивное построение структуры дерева через зарисовки отдельных деревьев, группы деревьев...
© cyberpedia.su 2017-2026 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!

