Ботачева Е. А.
Курс лекций по генетике микроорганизмов: учебное пособие для студентов
направления 020400 «Биология», бакалавриат, профиль подготовки
“Микробиология” / Е.А.Болтачева. – Киров: ПРИП ФГБОУ ВПО
«ВятГУ», 2014. – 60 с.
В издании излагаются основы генетики микроорганизмов в виде курса лекций. Курс формирует у обучающегося целостное представление о современной генетике вирусов и бактерий. Формирует представления о генетической детерминированности основных законов наследственности и изменчивости микроорганизмов, особенности структуры генома прокариот, способах передачи генетической информации, характеристиках мобильных генетических элементов и внехромосомных факторов наследственности прокариот, механизмах гибридизации бактерий (конъюгация, трансдукция, трансформация), а также прикладных аспектах (методах генетического конструирования микроорганизмов для использования их в качестве промышленных штаммов-продуцентов).
© ФГБОУ ВПО «ВятГУ» 2014
ISBN
Оглавление
1. Введение. Организация генома вирусов и фагов. 4
2. Организация генома клеток бактерий. 15
3. Бактериальные плазмиды.. 22
4. Механизмы гибридизации бактерий (трансформация, трансфекция) 29
5. Механизмы гибридизации бактерий (трансдукция) 36
6. Механизмы гибридизации бактерий (конъюгация) 43
7. Транспозирующиеся (мигрирующие) элементы прокариот. 52
8. Биотехнология микроорганизмов. 68
9. Вопросы для подготовки к экзамену по курсу «Генетика микроорганизмов» 78
ЛЕКЦИЯ 1
Введение. Организация генома вирусов и фагов
Вопросы:
1. Введение. Предмет и задачи лекционного курса.
2. Структура генома вирусов и фагов.
3. Типы генетического материала и механизмы его репликации у различных вирусов.
4. Бактериофаги.
5. Типы взаимодействия вирусов с клеткой-хозяином.
6. Гибридизация вирусов
7. Значение вирусов для эволюции организмов
Введение
Объектом изучения генетики является ряд поколений, возникающих в процессе размножения организмов. Мир микроорганизмов, включающий микроскопические грибы и одноклеточные водоросли, бактерии и вирусы, представляет исключительный интерес. По строению клеток и способу размножения высшие животные и растения не столь резко отличаются друг от друга, как микроорганизмы. Благодаря этому изучение генетики микроорганизмов раскрыло такие явления, о существовании которых нельзя было, и подозревать при генетическом исследовании высших организмов.
Значение генетики микроорганизмов, однако, заключается не только в обнаружении новых генетических явлений, но и главным образом в ее тесной связи с рядом других областей биологии. Генетика микроорганизмов не просто взяла на вооружение идеи и методы многих наук - общей генетики, микробиологии, биохимии, биофизики, она вместе с тем внесла существенный вклад в их развитие.
Обратимся, прежде всего, к общей генетике. До середины 40-х годов микроорганизмы не были ее популярными объектами. Представления о гене, о природе мутаций и, особенно, о репродукции гена и его функции носили в основном абстрактный характер. Но благодаря достижениям генетики микроорганизмов в 50-60-е годы удалось выяснить материальную основу
явлений наследственности и изменчивости. Решающее значение в этом отношении имели работы по трансформации у бактерий, воспроизведению фаговых частиц, химическому мутагенезу у бактерий и вирусов и биохимическим мутациям у грибов и бактерий. Это явилось фундаментом современной генетики и обусловило дальнейшие открытия в тех ее областях, где преобладал еще описательный метод.
Чрезвычайно велик вклад генетики микроорганизмов в молекулярную биологию. Если в 40-50-х годах речь шла о частных вопросах молекулярной биологии, то уже в 60-е годы выяснилось, что без генетики микроорганизмов невозможно не только решение, но даже и сама постановка актуальных и перспективных для современной молекулярной биологии проблем.
Что же касается микробиологии, то благодаря генетике она получила возможность обнаружения таких явлений в жизненном цикле микроорганизмов, которые ранее ускользали от внимания наблюдателей. Это, к примеру, открытия полового процесса и плазмид у бактерий, парасексуального процесса у несовершенных грибов, давшие ключ к пониманию эволюции микроорганизмов, их систематики, к решению ряда проблем в изучении популяций микроорганизмов и эволюции способов размножения у живых существ.
Открытие гибридизации вирусов - тоже весомый вклад генетики в вирусологию. Исследования, начатые на бактериофаге, в дальнейшем были перенесены на вирусы животных. Оказалось, что вирусы (например, вирус гриппа) обладают способностью к гибридизации. Этот факт не только представляет большой теоретический интерес, но и имеет важное значение для практической медицинской вирусологии: он указывает на пути происхождения некоторых штаммов при вирусных инфекциях, что особенно актуально для совсем недавно проявивших себя возбудителей птичьего и свиного гриппов.
Итак, все сказанное приводит к выводу, что генетика микроорганизмов в настоящее время занимает в биологии одну из ключевых позиций. Ее успехи являются условием развития смежных наук.
Задача настоящего лекционного курса состоит в изложении особенностей структуры генома прокариот, способов передачи генетической информации, характеристик мобильных генетических элементов и внехромосомных факторов наследственности прокариот, а также методов генетического конструирования микроорганизмов для использования их в качестве промышленных штаммов-продуцентов.
У различных вирусов
3.1 РНК-содержащие вирусы (РНК → РНК)
Геномы почти всех известных РНК-содержащих вирусов, реплицирующихся по данной схеме - это линейные молекулы, их удобно разделить на 2 группы.
Первая группа -это однонитевые геномы положительной полярности, т.е. с нуклеотидной последовательностью, соответствующей таковой у мРНК.

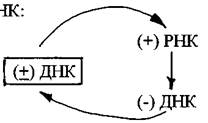
Такие геномы обозначают как (+) РНК. Цепь РНК такого типа может сразу после проникновения в клетку-хозяина обеспечивать синтез вирусных белков. Однонитевый (+) РНК-геном характерен для фага Qβ, вирусов табачной мозаики, полиомиелита, клещевого энцефалита. Вирусные (+) РНК-геномы кодируют несколько белков, среди которых РНК-зависимая РНК-полимераза (репликаза), способная синтезировать молекулы РНК без участия ДНК. С помощью этого фермента синтезируются сначала (-) нити РНК вируса, а затем репликаза осуществляет синтез (+) нити РНК. На заключительной стадии из накопившихся вирусных белков и (+) РНК формируются новые вирионы.
Вторая группа - это однонитевые геномы с негативной полярностью, т.е. (-) РНК-геномы. К вирусам с негативным РНК-геномом относятся вирусы гриппа, кори, бешенства. Поскольку (-) РНК не может выполнять функции мРНК, для образования «своих» мРНК вирус внедряет в клетку не только геном, но и фермент, умеющий снимать с этого генома комплиментарные копии по схеме:
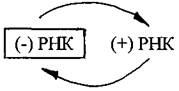
Этот вирусный фермент РНК-зависимая РНК-полимераза, синтезированная в предыдущем цикле размножения, упакован в вирионе в удобной для доставки в клетку форме. Инфекционный процесс начинается с того, что вирусный фермент копирует вирусный геном, образуя (+) РНК, которая выступает в качестве матрицы для синтеза вирусных белков, в том числе РНК-зависимой РНК-полимеразы, которая входит в состав образующихся вирионов.
ДНК-содержащие вирусы
Первая группа - вирусы с двуцепочечной ДНК, репликация которых осуществляется по схеме: ДНК→РНК→ДНК. Они получили название ретроидные вирусы. Представителями этой группы вирусов являются вирус гепатита В и вирус мозайки цветной капусты.
Репликация ДНК-генома этих вирусов осуществляется при посредстве промежуточных молекул РНК:
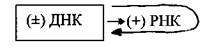
Молекулы (+) РНК образуются в результате транскрипции вирусных ДНК в клеточном ядре хозяйским ферментом ДНК-зависимой РНК-полимеразой. Транскрибируется только одна из нитей вирусной ДНК. Дальнейший синтез ДНК на РНК-матрице происходит в результате реакции, катализируемой обратной транскриптазой; сначала синтезируется (-) нить ДНК, а затем на вновь синтезированной (-) нити ДНК тот же фермент строит (+) нить. (+) РНК также используется для трансляции белков, необходимых для сборки вирусных частиц.
Вторая группа - вирусы с двухцепочечной ДНК, репликация которых осуществляется по схеме ДНК→ДНК. К этой группе относятся вирусы герпеса, оспы и др. В зараженной клетке ДНК-зависимая РНК-полимераза (фермент клетки-хозяина) транскрибирует с генома этих вирусов молекулы мРНК (т.е. (+) РНК), которые принимают участие в синтезе вирусных белков, а размножение вирусного генома осуществляет фермент ДНК-зависимая ДНК-полимераза:
Бактериофаги
Отмеченные особенности характерны для вирусов животных, растений и бактерий. Последние, называемые бактериофагами, наиболее детально изучены с генетической точки зрения. Связано это, прежде всего с легкостью их культивирования в лабораторных условиях.
Разные фаги обнаруживают специфичность действия - они действуют на бактерии лишь определенного вида, редко на близкие виды и часто - на отдельные штаммы данного вида. Фаги со специфическим приспособлением к тому или иному хозяину отличаются от других фагов морфологией, размерами, биохимическими и серологическими свойствами.
Биохимический анализ выявляет в фаговой частице два компонента -белок и нуклеиновую кислоту. Белок образует хвост фага и мембрану головки. При резком изменении осмотической силы среды мембрана может разрываться и ДНК освобождается.
Практически все известные фаги содержат ДНК. Молекулы у них состоят из двух комплементарных нитей. Каждая фаговая частица содержит лишь одну молекулу ДНК.
Поскольку размножение фагов происходит внутри бактериальной клетки, рассмотрим типичную картину взаимодействия фага и чувствительной бактерии.
Фаговые частицы адсорбируются на поверхности бактерии, прикрепляясь к ее оболочке своими хвостами. Зараженная бактерия через 15-45 мин лизируется, освобождая 100 и более зрелых частиц следующего поколения фага. Встречая чувствительные бактериальные клетки, они вновь заражают их, и весь цикл повторяется. Если размножение фага происходит в культуре в жидкой среде, то оно приводит к просветлению (из-за лизиса клеток) суспензии бактерий. Размножение фага на бактериях, растущих на плотной среде, выявляется в виде зон лизиса среди сплошного газона бактерий. Эти пятна обнаженного агара называются негативными колониями фага.
Гибридизация вирусов
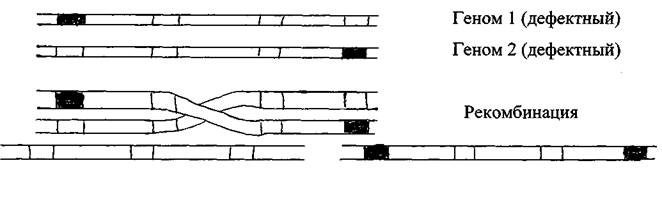
Заражение вирусами чувствительных к ним клеток носит множественный характер, т.е. в клетку может проникнуть несколько вирионов, обычно идентичных или близкородственных. В подобных ситуациях геномы вирусных частиц в динамике репродуктивных циклов могут взаимодействовать. Типами такого рода взаимодействия являются рекомбинациии и обмен фрагментами генома.
Рекомбинации могут иметь место у всех типов вирусов. В результате данных процессов даже два дефектных по разным сайтам вирусных генома могут образовать нормальный вирусный геном.
Геном 3 (нормальный) Геном 4 (дефектный)
Обмен фрагментами генома наблюдают у РНК-содержащих вирусов с сегментированным геномом, в частности, вируса гриппа. В отличие от рекомбинации суть процесса состоит в обмене крупных блоков наследственного материала. Этот феномен явился причиной появления таких патогенных для человека разновидностей вируса гриппа, как птичий грипп и свиной грипп. Формирование последних, скорее всего, происходило в организме промежуточных хозяев (не установлены) при одновременном проникновении в его клетку штаммов вируса обычного гриппа и непатогенного для человека штамма птичьего или свиного гриппа.
7 Значение вирусов для эволюции организмов
Вирусы играли и играют важную роль в эволюции тех организмов, которые они инфицируют. Многие вирусы часто вступают в рекомбинацию как друг с другом, так и с хромосомами клеток-хозяев; при этом они захватывают случайные фрагменты хромосом и переносят их в другие клетки или другие организмы. Кроме того, включившиеся в геном хозяина (интегрированные) копии вирусной ДНК (провирусы) становятся постоянными компонентами генома у большого числа организмов. Интегрированная вирусная ДНК часто видоизменяется так, что утрачивает способность образовывать полноценный вирус, но сохраняет способность кодировать белки, часть из которых оказывается полезной для клетки. Таким образом, вирусы, как и половой процесс, создают возможности для ускорения эволюции, открывая для нее такой путь, как смешение генофондов различных организмов.
ЛЕКЦИЯ 2
- Организация генома клеток бактерий
Вопросы:
1 Структура бактериальной хромосомы
2 Структуры, связанные с репликацией
3 Открытые рамки считывания
4 Минимальный размер генома прокариот
5 Оперонная организация генов прокариот
Открытые рамки считывания
В установленной нуклеотидной последовательности генома прокариот с помощью компьютерных программ выявлены открытые рамки считывания. Открытая рамка считывания (англ. ореn rеаding frame, ОRF) состоит из ряда триплетов, кодирующих аминокислоты; она не содержит каких-либо терминирующих кодонов; потенциально может транслироваться в белок. Средний размер их в прокариотических геномах соответствует примерно 300 аминокислотным остаткам. В настоящее время в хромосоме наиболее изученного микроорганизма - кишечной палочки, выявлено 4288 ОRF, однако функции 40% обнаруженных ОRF остаются неидентифицированными.
Сопоставление состава ОRF по функциям (также с помощью компьютерных программ) позволяет идентифицировать компоненты генома, общие для всех организмов, общие для данной группы видов и уникальные,
специфичные только для данного организма. Функциональная классификация генов и генных продуктов является основной для сравнения геномов и выявления сходства и различия их метаболизма.
ЛЕКЦИЯЗ
Бактериальные плазмиды
Вопросы:
1 Структура и репликация бактериальных плазмид
2 Генетическая организация плазмид
3 Несовместимость плазмид
Несовместимость плазмид
Плазмиды эгоистичны. Завладев бактериальной клеткой, плазмида старается предотвратить проникновение и размножение в ней любой другой плазмиды того же типа. Для этого используются два независимых механизма: поверхностное исключение и плазмидная несовместимость.
Поверхностное исключение определяет неспособность плазмид внедряться в клетки, уже несущие другую плазмиду того же типа. Эффект проявляется на поверхности бактерии и характеризует систему, используемую при переносе плазмид в процессе конъюгации.
Установлено 4 типа систем поверхностного исключения, фенотипическое выражение которых характеризуется двумя основными проявлениями. Первое из них заключается в том, что при формировании конъюгационных пар «донор-реципиент» блокируется фиксация кончика половой ворсинки (пили) на определенном специфическом рецепторе поверхности клетки-реципиента, что делает невозможным конъюгационную передачу плазмидной ДНК. Подобный эффект имеет место при проявлении механизма поверхностного исключения, детерминированного плазмидой К100. Во втором случае поверхностное исключение достигается путем подавления выхода ДНК из клетки бактерии-донора, что особенно характерно для полового фактора кишечной палочки.
При передаче плазмид между клетками бактерий, имеющими различные типы поверхностного исключения, ограничений в этом процессе практически не наблюдается.
В том случае, если в бактериальную клетку, уже несущую плазмидную ДНК, тем или иным способом внедряется дополнительная чужеродная плазмида, могут иметь место два исхода. При одном из них обе плазмиды могут нормально реплицироваться и сегрегировать в дочерние клетки. В другом случае наблюдается подавление репликации одной плазмиды в присутствии другой и в процессе размножения клетки наследуют только одну из двух плазмид либо избирательно, либо случайно. Данное явление получило название «плазмидная несовместимость».
Несовместимость связана с жесткой регуляцией контроля числа копий плазмид. Данный контроль достигается путем синтеза репрессора, определяющего число точек начала репликации. Введение новой точки начала репликации при внедрении в клетку другой, сходной плазмиды имитирует начало репликации резидентной плазмиды; теперь присутствуют две точки начала репликации. В результате любая дальнейшая репликация плазмид предотвращается до тех пор, пока они не разойдутся в разные клетки или одна из плазмид не элиминируется.
Явление несовместимости используют для классификации плазмид. Применяя несовместимость в качестве критерия, многие из известных плазмид были распределены на ряд групп несовместимости. Последние определяются как группы плазмид, члены которых не способны сосуществовать в одной и той же бактериальной клетке. К настоящему времени известно более 20 групп несовместимости, в состав которых входят от одной до нескольких десятков плазмид. Плазмиды различных групп несовместимости детерминируют образование разных типов половых пилей,а несущие их клетки характеризуются разной специфической фагочувствительностью.
В то же время присутствие в бактериальной клетке плазмиды, относящейся к одной группе несовместимости, не влияет на выживаемость плазмиды, принадлежащей к другой группе несовместимости.
ЛЕКЦИЯ4
Вопросы
1 Трансформация, общая характеристика явления
2 Стадии трансформации
3 Трансфекция
Стадии трансформации
Процесс трансформации протекает в несколько стадий. Вначале ДНК адсорбируется на клеточной стенке, затем проникает внутрь и, наконец, рекомбинирует с клеточной хромосомной ДНК.
Итак, первый этап - связывание экзогенной ДНК с поверхностью компетентных клеток. Адсорбированная на этом этапе ДНК не отделяется от клеток при их отмывании. Адсорбция ДНК происходит на специальных структурах, обеспечивающих её проникновение через клеточную стенку и плазматическую мембрану. В частности, на поверхности компетентной клетки НаеторЬПш тйиепхае образуются везикулярные структуры и меняется липидный и белковый состав клеточной стенки.
Связь между поверхностными структурами и ДНК нековалентная - при обработке детергентами или фенолом ДНК отделяется от клетки. Природным субстратом для связывания служит только двухцепочечная молекула ДНК.
Следующий этап - фрагментация, при которой обе цепи связанной ДНК подвергаются разрезанию, которое происходит в случайных точках, но приводит к образованию фрагментов с определенным распределением по размерам. В связанном состоянии ДНК остается чувствительной к воздействию ДНКазы, образуя на поверхности клетки вытянутую структуру. Средняя величина образующихся при разрезании фрагментов составляет примерно 19 т.п.н.
Третий этап - поглощение трансформирующей ДНК. На этой стадии связанная с клеткой-реципиентом ДНК в течение 1-2 минут полностью теряет чувствительность к ДНКазе. Считается, что это обусловлено либо транспортом ДНК через мембрану, либо проникновением её в участки, недоступные для действия нуклеаз. ДНК поглощается как линейная молекула со скоростью 50 - 200 п.н./сек. В процессе поглощения одна цепь трансформирующей ДНК подвергается деградации под действием нуклеаз и внутрь бактериальной клетки проникает только одноцепочечная молекула ДНК в виде составляющих её фрагментов.
Наиболее детально этот этап трансформации описан у НаеторЫ1ш тйиепгае. Поглощение ДНК у данного микроорганизма носит характер высокоизбирательного процесса в отношении гомологичной ДНК. Распознавание гомологичной и гетерологичной ДНК обеспечивают присутствующие в ДНК этого микроба «сайты поглощения», состоящие из 11 п.н. и повторяющиеся через интервалы длиной 4 т.п.н. В этом процессе, а также в поглощении экзогенной ДНК и её защите от нуклеаз также участвуют везикулярные мембранные выросты, называемые трансфор-мосомами. Они локализованы преимущественно в точках контакта плазматической и наружной мембран. Добавление гомологичной ДНК вызывает переход трансформосом во внутреннее пространство клетки с потерей чувствительности ДНК к нуклеазам. После проникновения в цитоплазму происходит высвобождение ДНК из трансформосом.
Параллельно этому у НаеторЬПш тйиепгае возрастает способность к рекомбинации - в хромосомной ДНК возникают одноцепочечные бреши и «хвосты», т.е. она как бы подготавливается для включения экзогенной ДНК.
Завершающий этап трансформации - интеграция ДНК, которая осуществляется по принципу «разрыв-воссоединение». В ДНК клетки-реципиента включается лишь одна цепь трансформирующей ДНК, которая спаривается с гомологичным участком ДНК в реципиентной клетке, с цепью, являющейся комплементарной ей. Продукт рекомбинации представляет собой спаренный трехцепочечный гетеродуплекс из донорной и реципиентной молекул ДНК. Поскольку донорная ДНК ранее была подвергнута фрагментации на поверхности клетки, в процессе её интеграции происходит ряд независимых рекомбинационных событий в различных участках реципиентной ДНК.
Затем в результате двойного кроссинговера между однонитчатой донорной ДНК и двунитчатой ДНК реципиента происходит образование рекомбинантной хромосомы реципиента. При этом в участке ДНК, ограниченном сайтами кроссинговера, одна нить ДНК имеет реципиентный сегмент, другая - донорный. Уже после первого раунда репликации ДНК образуются два типа клеток: исходные и трансформированные, которые несут ДНК донора.
Таким образом, первые стадии трансформации - связывание экзогенной ДНК, её фрагментация, поглощение и деградация одной цепи -осуществляются с равной эффективностью независимо от её гомологии с ДНК реципиента. Эффективность последней стадии трансформации, а именно интеграция чужеродной ДНК, напрямую связана со степенью гомологии между донорной и реципиентной ДНК.
В отличие от вышеуказанных грамположительных микроорганизмов, у грамотрицательных колиформных бактерий состояния высокоэффективной природной компетентности в том виде, в каком оно было рассмотрено выше, вообще не существует. По-видимому, трансформация не служит у этих организмов важным механизмом обмена генетической информацией. Однако в связи с тем, что кишечная палочка имеет важное значение как модель для генетических исследований, поиску способов её трансформации были посвящены специальные работы.
Как удалось установить, обработка клеток кишечной палочки раствором СаС\2 или смеси других солей на холоду делает их компетентными для поглощения экзогенной ДНК в течение короткого периода нагревания. Другим способом создания компетентности у данного вида бактерий (и не только у них) является введение ДНК с помощью электропорации, при которой клетки подвергаются воздействию высоковольтного электрического заряда, что обеспечивает проникновение ДНК через клеточную мембрану. В отличие от существующего в природе высокоэффективного механизма трансформации с превращением экзогенной ДНК в линейную одноцепочечную форму, при искусственной компетентности клетки кишечной палочки поглощают кольцевую двухцепочечную ДНК.
Трансформация служит хорошим инструментом для картирования хромосом, поскольку трансформированные клетки включают различные фрагменты ДНК. Определение частоты одновременного приобретения двух заданных характеристик (чем ближе друг к другу расположены гены, тем более вероятно, что они оба включатся в один и тот же участок ДНК) даёт информацию о взаиморасположении соответствующих генов в хромосоме. Таким образом, трансформацию активно используют для определения порядка расположения генов, расстояний между ними в молекуле ДНК и построения генетических карт.
Трансфекция
Частным случаем трансформации является трансфекция. Данное явление впервые было описано в 1964 году на примере ВасШш зиЫШз. Его сущность заключается в продуктивной инфекции бактерий данного вида ДНК, выделенной из лизирующих их фагов.
Как и в случае классической трансформации, чувствительными к трансфекции являются лишь те клетки, которые компетентны к восприятию трансформирующей ДНК.
Явление трансфекции также установлено при обработке клеток кишечной палочки ДНК, выделенной из разных кишечных фагов, а также в случае инфицирования растений табака инфекционной РНК, выделенной из вируса мозаичной болезни табака. Ряду исследователей удалось наблюдать формирование вируса оспы после инфицирования компетентных клеток млекопитающих ДНК, выделенной из оспенного вируса.
Механизм трансфекции зависит от рекомбинационной системы клеток-реципиентов и рекомбинационный механизм трансфекции отличается от рекомбинационного механизма трансформации. В частности, трансфекция
ВасШиз зиЫШз ДНК из фага Н1 требует объединения 4-5 молекул фаговой ДНК. Механизм компетентности клеток к трансформации и трансфекции одинаковый.
ЛЕКЦИЯ5
Вопросы
1 Общая характеристика явления
2 Общая (неспецифическая) трансдукция
3 Специфическая (ограниченная) трансдукция
4 Абортивная трансдукция
Абортивная трансдукция
Устойчивое наследование перенесенных в результате трансдукции генов возможно только при их включении в геном бактериальной клетки. Если в силу различных причин ген не может быть включенным в хромосому, он присутствует в отдельных клетках бактериальной популяции в виде единственной копии. Такой вид переноса генов называется абортивной трансдукцией.
Перенесенная ДНК в этом случае транскрибируется (происходит синтез продуктов), но не реплицируется. При посеве на агаровую среду, селективную для клеток, содержащих перенесенный ген, образуются микроколонии, содержащие примерно 104 клеток. Их рост происходит в результате того, часть дочерних клеток-реципиентов ещё содержат продукт абортивно трансдуцированного гена и, благодаря этому, они выживают в селективных условиях. Дальнейшее деление клеток и оборот продукта приводят к его исчерпанию, и рост таких клеток после нескольких генераций прекращается, то есть трансдуцированный ген теряется в потомстве.
ЛЕКЦИЯ 6
Вопросы
1 Общая характеристика явления.
2 Механизм конъюгации у грамотрицательных бактерий.
3 Конъюгация у грамположительных бактерий
ЛЕКЦИЯ 7
Вопросы
1. Общая характеристика
2 18-элементы
3 Транспозоны
4 Конъюгагивные транспозоны
5 18-элементы и транспозоны в плазмидах
6 Значение транспозирующихся генетических элементов для эволюции микроорганизмов
Общая характеристика
Согласно представлениям, сложившимся в рамках классической (формальной) генетики, геномы различных организмов характеризуются значительной стабильностью и подвержены лишь очень медленным эволюционным преобразованиям. Эти представления существенно изменились в связи с открытием транспозирующихся (мигрирующих) генетических элементов (фрагментов ДНК), которые могут перемещаться как в пределах одного генома, так и из одного генома в другой. Способность к транспозиции закодирована в нуклеотидных последовательностях самих мигрирующих фрагментов ДНК и связана с возможностью их включения (интеграции) в различные места (сайты) хромосом и внехромосомных молекул ДНК и
выхода («вырезания») из этих сайтов. Такие мобильные структуры были обнаружены в геномах как прокариот, так и эукариот.
Транспозирующиеся элементы первоначально были открыты при обнаружении вставок (инсерций) нового материала в пределах бактериальных оперонов. Такие вставки локализовались внутри гена и предотвращали его транскрипцию и (или) трансляцию. Позднее было установлено, что транспозирующиеся элементы могут непосредственно влиять на геном хозяина, поскольку способны внедрятся не только внутрь генов, но и в близко прилегающие участки, нарушая генную структуру или подчиняя экспрессию этих генов новым регуляторным элементам. Рекомбинация между двумя копиями одного и того же мобильного элемента может приводить к делециям, дупликациям, амплификациям, инверсиям и транспозициям сегментов генома. Мобильные элементы вызывают многие хромосомные мутации и обуславливают их высокую частоту.
У бактерий транспозирующиеся генетические элементы представлены сегментами ДНК двух типов - инсерционными (вставочными) последовательностями или 18-элементами и транспозонами. Те и другие могут перемещаться в границах одной генетической структуры (бактериальной хромосомы, плазмиды) либо из одной структуры в другую например, из хромосомы в плазмиду, из плазмиды в хромосому, из оной плазмиды в другую плазмиду, из плазмиды бактерии одного вида в плазмиду либо хромосому бактерии другого вида и т.д.).
Элементы
Первая группа транспозирующихся генетических элементов получила название инсерционных последовательностей. Каждый тип инсерционной последовательности обозначается префиксом 18, далее следует номер, соответствующий именно этому типу, например, 181, 182, 183 и т.д. 18-элементы были открыты Старлинжером, Сердлером и Шапиро в начале 70-х годов. Они являются нормальными компонентами бактериальной хромосомы и плазмид. В частности, один стандартный штамм кишечной палочки
содержит 5-10 копий 181, 4-8 копий 182, 10-12 копий 1810 ряд других. Для обозначения инсерции в определенном сайте используется двоеточие; так, обозначение лямбда:: 181 показывает, что 181-элемент внедрен в геном фага лямбда.
Ген, кодирующий транспозазу-белок, необходимый для перемещения
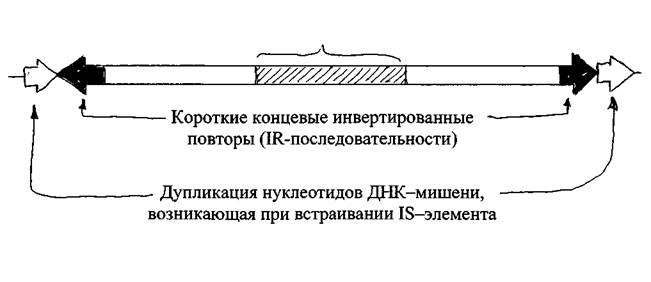
Рис. 1 18 - элемент кишечной палочки
Строение типичного 18-элемента представлено на рис. 1. 18-элемен-ты - это небольшие участки ДНК, размеры которых обычно варьируют в пределах порядка от 750 до 1550 пар нуклеотидов. На концах (на флангах) этих участков имеются одинаковые либо очень сходные короткие нуклеотидные последовательности (20-40 пар нуклеотидов), расположенные в обратном порядке (инвертированные повторы или 1К-последовательности). Считают, что ^-последовательности играют важную роль в процессах перемещения транспозирующихся элементов. В центральной части все 18-элементы содержат ген, кодирующий синтез белка, названного транспозазой.
В процессе перемещения 18-элементов происходит точное копирование уже встроенного элемента, затем старая копия остается на месте, а вновь синтезированная внедряется в новый сайт. Репликация новой копии происходит с использованием энзимов репликационной машины клетки-хозяина. Процесс транспозиции осуществляется с использованием транспозазы, которая распознает 1К-последовательности, где и инициируется транспозиция.
Внедрение 18-элемента в новый локус сопровождается дупликацией (прямой повтор) последовательностей из нескольких нуклеотидов (5-9 п.н.) данного локуса. Число дуплицированных нуклеотидов специфично для каждого 18-элемента.
Разные 18-элементы являются типичными обитателями бактерий и обычно выявляются в количествах, превышающих одну копию на клетку. Наличие идентичных 18-элементов в разных бактериальных структурах (хромосомах, плазмидах) создает основу для их взаимодействия по механизму сайт-специфической рекомбинации, что может приводить к объединению (коинтеграции) таких структур, либо к генетическому обмену между ними. В отличие от генетической рекомбинации общего типа, требующей наличия значительных областей гомологии ДНК у двух взаимодействующих родственных структур, сайт-специфическая рекомбинация основана на том, что соответствующий фермент (транспозаза) узнает небольшие специфические последовательности в двух местах (сайтах) одной, либо разных молекул ДНК, где и происходит процесс рекомбинации. Следовательно, такая рекомбинация может приводить к генетическому обмену между неродственными структурами (хромосомами, плазмидами).
В целом, 18-элементы представляют собой простейший класс транспозонов; их генетические функции связаны только со способностью к транспозиции.
Транспозоны
Транспозонами (Тп-элементами) называют сегменты ДНК, обладающие такими же свойствами, что и 18-элементы, но содержащие также, гены, не имеющие непосредственного отношения к транспозиции
л 4
' Ь
(гены устойчивости к антибиотикам, гены токсинов, гены дополнительных ферментов клеточного метаболизма и т.д.). В общем, для транспозонов характерны те же гены, которые имеются в плазмидах. Более того, нередко присутствие в составе плазмиды того или иного гена обусловлено наличием в последовательности плазмидной ДНК соответствующего транспозона.
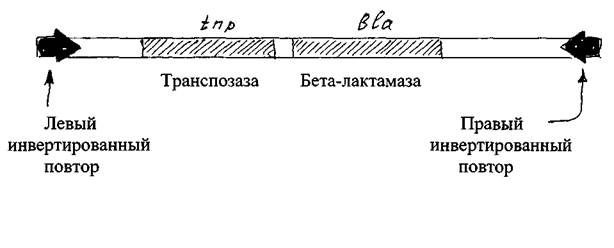
Известны два типа транспозонов прокариот: простые и сложные. Простые транспозоны имеют центральный район, содержащий гены, например, ген синтеза бета-лактамазы, обуславливающей устойчивость к бета-лактамным антибиотикам, и ген, кодирующий синтез транспозазы. Подобно 18-элементам, простые транспозоны всегда имеют на своих концах инвертированные повторы, необходимые для транспозиций (рис. 2).
А
Рис. 2 Структура простого транспозона ТпЗ
Сложные транспозоны также имеют центральный район, содержащий гены. На обоих концах транспозона расположены 18-элементы, которые в пределах одного транспозона принадлежат к одному типу и называются «левый» и «правый». Таким образом, центральный район сложного транспозона фланктрован инвертированными 18-элементами, которые, в свою очередь, имеют свои собственные терминальные инвертированные повторы (рис. 3).
^
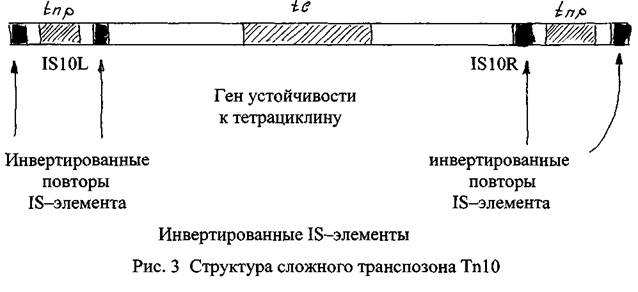
Другое важное отличие сложных транспозонов от простых заключается в том, в большинстве случаев в центральном районе сложных транспозонов отсутствует ген, кодирующий синтез транспозазы, а продукция данного фермента осуществляется с помощью генов, дислоцированных в составе фланкирующих транспозон 18-элементов.
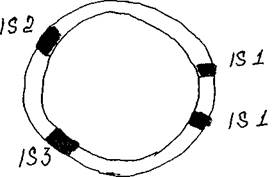
Рис. 4 Образование транспозона близкорасположенными 18-элементами Важно отметить, что часто два близкорасположенных 18-элемента способны перемещаться вместе, одновременно перенося заключенный между ними сегмент ДНК. Таким образом, транспозон могут образовать два расположенных рядом 18-элемента (рис. 4).
Специфические концевые повторы 18-элементов и транспозонов играют ключевую роль в процессах транспозиции, поскольку они распознаются ферментами, осуществляющими сайт-специфическую рекомбинацию. При транспозиции транспозонов работает механизм, как правило, не связанный с процессом полного «вырезания» всей структуры из одного сайта и её перемещения в другой сайт. При этом происходит удвоение (репликация) генетического элеме