Нервная система позвоночных закладывается в виде сплошной нервной трубки, которая в процессе онто— и филогенеза дифференцируется на различные отделы и является также источником периферических симпатических, парасимпатических и метасимпатических нервных узлов. У наиболее древних хордовых (бесчерепных) головной мозг отсутствует, и нервная трубка представлена в малодифференцированном состоянии.
Согласно представлениям Л. А. Орбели, С. Херрика, А. И. Карамяна, этот критический этап развития центральной нервной системы обозначается как спинальный. Нервная трубка современного бесчерепного (ланцетника), как и спинной мозг более высокоорганизованных позвоночных, имеет метамерное строение и состоит из 62—64 сегментов, в центре которых проходит спинномозговой канал. От каждого сегмента отходят брюшные (двигательные) и спинные (чувствительные) корешки, которые не образуют смешанных нервов, а идут в виде отдельных стволов. В головных и хвостовых отделах нервной трубки локализованы гигантские клетки Родэ, толстые аксоны которых образуют проводниковый аппарат. С клетками Родэ связаны светочувствительные глазки Гесса, возбуждение которых вызывает отрицательный фототаксис
В головной части нервной трубки ланцетника находятся крупные ганглиозные клетки Овсянникова, имеющие синаптические контакты с биполярными чувствительными клетками обонятельной ямки. В последнее время в головной части нервной трубки идентифицированы нейросекреторные клетки, напоминающие гипофизарную систему высших позвоночных. Однако анализ восприятия и простых форм обучения ланцетника показывает, что на данном этапе развития ЦНС функционирует по принципу эквипотенциальности, и утверждение о специфике головного отдела нервной трубки не имеет достаточных оснований.
В ходе дальнейшей эволюции наблюдается перемещение некоторых функций и систем интеграции из спинного мозга в головной — процесс энцефализации, который был рассмотрен выше на примере беспозвоночных животных. В период филогенетического развития от уровня бесчерепных до уровня круглоротых формируется головной мозг как надстройка над системами дистантной рецепции.
Исследование ЦНС современных круглоротых показывает, что их головной мозг в зачаточном состоянии содержит все основные структурные элементы. Развитие вестибулолатеральной системы, связанной с полукружными каналами и рецепторами боковой линии, возникновение ядер блуждающего нерва и дыхательного центра создают основу для формирования заднего мозга.
Развитие дистантной зрительной рецепции дает толчок к закладке среднего мозга. На дорсальной поверхности нервной трубки развивается зрительный рефлекторный центр — крыша среднего мозга, куда приходят волокна зрительного нерва. И наконец, развитие обонятельных рецепторов способствует формированию переднего, или конечного, мозга, к которому примыкает слаборазвитый промежуточный мозг.
Указанная выше направленность процесса энцефализации согласуется с ходом онтогенетического развития мозга у круглоротых. В процессе эмбриогенеза головные отделы нервной трубки дают начало трем мозговым пузырям. Из переднего пузыря формируется конечный и промежуточный мозг, средний пузырь дифференцируется в средний мозг, а из заднего пузыря образуются продолговатый мозг и мозжечок. Сходный план онтогенетического развития мозга сохраняется и у других классов позвоночных.
Нейрофизиологические исследования мозга круглоротых показывают, что его главный интегративный уровень сосредоточен в среднем и продолговатом мозгу, т. е. на данном этапе развития ЦНС доминирует бульбомезенцефальная система интеграции, пришедшая на смену спинальной.
Передний мозг круглоротых длительное время считали чисто обонятельным. Однако исследования недавнего времени показали, что обонятельные входы в передний мозг не являются единственными, а дополняются сенсорными входами других модальностей. Очевидно, уже на ранних этапах филогенеза позвоночных передний мозг начинает участвовать в переработке информации и управлении поведением. Вместе с тем энцефализация как магистральное направление развития мозга не исключает эволюционных преобразований в спинном мозгу круглоротых. В отличие от бесчерепных нейроны кожной чувствительности выделяются из спинного мозга и концентрируются в спинномозговой ганглий. Наблюдается совершенствование проводниковой части спинного мозга. Проводящие волокна боковых столбов имеют контакты с мощной дендритной сетью мотонейронов. Формируются нисходящие связи головного мозга со спинным через мюллеровские волокна — гигантские аксоны клеток, лежащих в среднем и продолговатом мозгу.
Появление более сложных форм двигательного поведения у позвоночных сопряжено с совершенствованием организации спинного мозга. Так, например, переход от стереотипных ундулирующих движений круглоротых к локомоции с помощью плавников у хрящевых рыб (акулы, скаты) связан с разделением кожной и мышечно—суставной (проприоцептивной) чувствительности. В спинальных
ганглиях появляются специализированные нейроны для выполнения этих функций.В эфферентной части спинного мозга хрящевых рыб также наблюдаются прогрессивные преобразования. Укорачивается путь моторных аксонов внутри спинного мозга, происходит дальнейшая дифференциация его проводящих путей. Восходящие пути боковых столбов у хрящевых рыб доходят до продолговатого мозга и мозжечка. Вместе с тем восходящие пути задних столбов спинного мозга еще не дифференцированы и состоят из коротких звеньев.
Нисходящие пути спинного мозга у хрящевых рыб представлены развитым ретикулярно—спинномозговым путем и путями, соединяющими вестибулолатеральную систему и мозжечок со спинным мозгом (преддверно—спинномозговой и мозжечково—спинномозговой пути).
Одновременно в продолговатом мозгу наблюдается усложнение системы ядер вестибулолатеральной зоны. Этот процесс сопряжен с дальнейшей дифференциацией органов боковой линии и с появлением в лабиринте третьего (наружного) полукружного канала в дополнение к переднему и заднему.
Развитие общей двигательной координации у хрящевых связано с интенсивным развитием мозжечка. Массивный мозжечок акулы имеет двусторонние связи со спинным, продолговатым мозгом и покрышкой среднего мозга. Функционально он разделяется на части: старый мозжечок (археоцеребеллум), связанный с вестибулолатеральной системой, и древний мозжечок (палеоцеребеллум), включенный в систему анализа проприоцептивной чувствительности. Существенным моментом структурной организации мозжечка хрящевых рыб является его многослойность. В сером веществе мозжечка акулы идентифицированы молекулярный слой, слой клеток Пуркинье и зернистый слой.
Другой многослойной структурой стволовой части мозга хрящевых рыб является крыша среднего мозга, куда подходят афференты различных модальностей (зрительные, соматические). Сама морфологическая организация среднего мозга свидетельствует о его важной роли в интегративных процессах на данном уровне филогенетического развития.
В промежуточном мозгу хрящевых рыб происходит дифференциация гипоталамуса, который является наиболее древним образованием этой части мозга. Гипоталамус имеет связи с конечным мозгом. Сам конечный мозг разрастается и состоит из обонятельных луковиц и парных полушарий. В полушариях у акул находятся зачатки старой коры (археокортекса) и древней коры (палеокортекса).
Палеокортекс, тесно связанный с обонятельными луковицами, служит главным образом для восприятия обонятельных стимулов. Археокортекс, или гиппокампальная кора, предназначен для более сложной обработки обонятельной информации. Вместе с тем электрофизиологические исследования показали, что обонятельные проекции занимают только часть полушарий переднего мозга. Кроме обонятельной здесь обнаружено представительство зрительной и соматической сенсорных систем. Очевидно, старая и древняя кора может участвовать в регуляции поисковых, пищевых, половых и оборонительных рефлексов у хрящевых рыб, многие из которых являются активными хищниками.
Таким образом, у хрящевых рыб складываются основные черты ихтиопсидного типа организации мозга. Его отличительной чертой является присутствие надсегментарного аппарата интеграции, координирующего работу моторных центров и организующего поведение. Эти интегративные функции осуществляют средний мозг и мозжечок, что позволяет говорить о мезэнцефалоцеребеллярной системе интеграции на данном этапе филогенетического развития нервной системы. Конечный мозг участвует в регуляции функций нижележащих отделов.
Переход позвоночных от водного к наземному образу жизни связан с целым рядом перестроек в ЦНС. Так, например, у амфибий в спинном мозгу появляется два утолщения, соответствующих тазовому и грудному поясам конечностей. В спинальных ганглиях вместо биполярных чувствительных нейронов сосредоточиваются униполярные с Т—образно ветвящимся отростком, обеспечивающие высокую скорость проведения возбуждения без участия клеточного тела. На периферии в коже земноводных формируются специализированные рецепторы и рецепторные поля, обеспечивающие дискриминационную чувствительность. В мозговом стволе также происходят структурные изменения в связи с перераспределением функциональной значимости различных отделов. В продолговатом мозгу наблюдаются редукция ядер боковой линии и формирование улиткового слухового ядра, осуществляющего анализ информации от примитивного органа слуха
По сравнению с рыбами у амфибий, имеющих довольно стереотипную локомоцию, наблюдается значительная редукция объема мозжечка. Средний мозг, так же как и у рыб, представляет собой многослойную структуру, в которой наряду с ростральными холмиками — ведущим отделом интеграции зрительного анализатора — имеются дополнительные бугорки — предшественники каудальных холмиков пластинки крыши.
Наиболее существенные в эволюционном плане изменения происходят в промежуточном мозгу амфибий. Здесь обособляется таламус (зрительный бугор), дифференцируются структурированные ядра (наружное коленчатое тело) и восходящие пути, связывающие зрительный бугор с корой (таламокортикальный путь).
В полушариях переднего мозга происходит дальнейшая дифференциация зачатков старой и древней коры. В старой коре (археокортексе) обнаруживаются звездчатые и пирамидные клетки. В промежутке между старой и древней корой появляется полоска плаща, которая является предтечей новой коры (неокортекса).
В целом развитие переднего мозга создает предпосылки для перехода от свойственной рыбам мезэнцефалоцеребреллярной системы интеграции к диэнцефалотелэнцефалъной, где ведущим отделом становится передний мозг, а таламус промежуточного мозга превращается в коллектор всех афферентных сигналов. В полной мере эта система интеграции представлена в зауропсидном типе мозга у рептилий и знаменует собой следующий этап морфофункциональной эволюции мозга.
Развитие таламокортикальной системы связей у рептилий приводит к формированию новых проводящих путей, как бы подтягивающихся к филогенетически молодым формациям мозга.
В боковых столбах спинного мозга рептилий обособляется восходящий спинно—таламический путь, который проводит к головному мозгу информацию о температурной и болевой чувствительности. Здесь же в боковых столбах формируется новый нисходящий путь — красно—ядерно—спинномозговой (Монакова). Он связывает мотонейроны спинного мозга с красным ядром среднего мозга, которое включено в древнюю экстрапирамидную систему двигательной регуляции. Эта многозвенная система объединяет влияние переднего мозга, мозжечка, ретикулярной формации ствола, ядер вестибулярного комплекса и координирует двигательную активность. У рептилий, как истинно наземных животных, возрастает роль зрительной и акустической информации, возникает необходимость сопоставления этой информации с обонятельной и вкусовой. В соответствии с этими биологическими изменениями в стволовой части мозга рептилий происходит целый ряд структурных изменений. В продолговатом мозгу дифференцируются слуховые ядра, помимо улиткового ядра появляется угловое, связанное со средним мозгом. В среднем мозгу двухолмие преобразуется в четверохолмие, в ростральных холмах которого находятся акустические центры.
Наблюдается дальнейшая дифференциация связей крыши среднего мозга с таламусом, который является как бы преддверия входа в кору всех восходящих сенсорных путей. В самом таламусе происходит дальнейшее обособление ядерных структур и установление между ними специализированных связей.
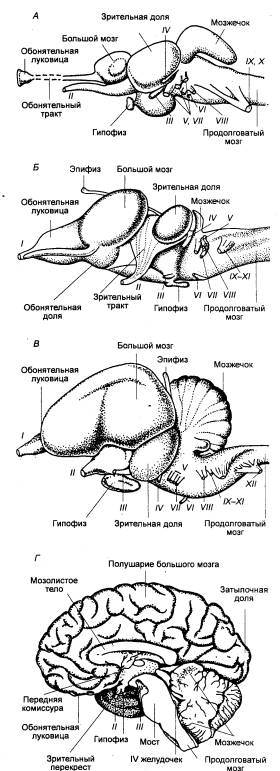
Рис. 3.1 Головной мозг позвоночных животных

девиации. При этом рекапитуляция (повторение предковых признаков) наблюдается только до средних стадий эмбриогенеза, а затем развитие идет по новому пути.
Архаллаксис — изменения начальных стадий эмбриогенеза или изменения самих зачатков органа. Этим путем идет развитие волоса млекопитающих — производного кожи — без повторения предковых признаков. Архаллаксисы вызывают с самого начала коренную перестройку в развитии органа. Они могут быть причиной нарушения функции органа и его связей с другими частями организма, что может привести к смерти. Очевидно, поэтому в филогенезе они встречаются реже, чем другие филэмбриогенезы. При архаллаксисе не наблюдается палингенезов и рекапитуляции и поэтому положения биогенетического закона здесь неприемлемы.
Следует отметить, что разные типы филэмбриогенезов не обособлены, они связаны и имеют взаимопереходы. Филэмбриогенезы характерны л для растений. Они возникают на разных стадиях развития и могут быть положительными (возникновение новых признаков) и отрицательными (выпадение, утрата старого признака).
Если биогенетический закон фиксирует внимание на зависимости онтогенеза от филогенеза (Ф→О), то теория филэмбриогенезов показывает, что и изменения в онтогенезе влияют на филогенез (Ф↔О) — онтогенетическая обусловленность филогенеза.








