Хромосомный уровень организации наследственного материала характеризуется особенностями морфологии и функций хромосом. Роль хромосом в передаче наследственной информации была доказана благодаря:
1) открытию хромосомного определения пола,
2) установлению групп сцепления генов, соответствующих числу хромосом,
3) построению генетических и цитологических карт хромосом.
У ДНК-содержащих вирусов, бактерий, сине-зеленых водорослей, а также в самореплицирующихся органеллах клеток эукариот (пластиды, митохондрии, кинетопласты и др.) наследственный материал представлен единственной хромосомой, которая представляет собой голую двуспиральную молекулу ДНК. Молекула эта у некоторых форм линейна, но у большинства образует кольцо, которое обычно перекручено и имеет суперспирализованный вид. Длина молекул ДНК-содержащих вирусов, прокариот и клеточных органелл составляет: от 5 до 100 мк. У наиболее мелких вирусов - от 0,4 до 1 мк, а у бактерий - 1000-2000 мк.
У большинства РНК-содержащих вирусов хромосома представлена голой однонитевой молекулой РНК, например у ВИЧ. Однако известны вирусы, у которых хромосома образована двунитевой молекулой РНК. Размеры хромосом РНК-содержащих вирусов меньше, чем у ДНК-содержащих вирусов.
В ДНК вирусов закодирована информация обо всех его структурных белках. Многие вирусы содержат гены специфических ферментов, контролирующих репликацию фермента клетки-хозяина. Мелкие вирусы содержат только 3 гена, которые кодируют А-белок, репликазу, белок оболочки. Гены вирусов могут существовать в виде фрагментов ДНК, разделенных генетически инертными нуклеотидными последовательностями, которые в момент работы генов "вырезаются" и целостность генетической информации восстанавливается.
Транскрипция и репликация генетической информации осуществляется с участием ферментов клетки-хозяина.
Хромосомы прокариот представлены голой кольцевой молекулой ДНК. Прокариоты содержат только по одной хромосоме и являются гаплоидами. Молекулярная масса ДНК прокариот соответствует примерно 2000 структурных генов, длиной около 1 500 пар азотистых оснований. Гены располагаются линейно и несут информацию о структуре 3-х – 4,5 тысяч различных белков.
Хромосомы эукариот, в отличие от хромосом прокариот, построены из нуклеопротеида, главными компонентами которого являются ДНК и два типа белков - гистоновых (основных) и негистоновых (кислых) белков. Установлено, что в хромосомах эукариот (за исключением политенных хромосом) имеется лишь одна непрерывная нить ДНК, представляющая единую гигантскую двуспиральную молекулу, состоящую из сотен миллионов пар нуклеотидов. Длина ДНК в хромосоме может достигать нескольких сантиметров. Подтвердилось блестящее представление Н.К. Кольцова, который писал ещё в 30-х годах: «В основе каждой хромосомы лежит тончайшая нить, которая представляет собой спиральный ряд огромных органических молекул - генов. Возможно, что эта спираль является одной гигантской молекулой». Молекулярная организация хромосомы была рассмотрена нами ранее. В метафазе хромосомы, состоящие из двух сильно спирализованных хроматид, хорошо заметны, но гены в них остаются неактивными на протяжении всего митоза. После окончания митоза происходит деспирализация хромосом. В интерфазном ядре хромосома состоит из сильно растянутой хроматиды. Из-за небольшой толщины (25 мк) они не видны в оптический микроскоп, но хорошо видны в электронном микроскопе и не теряют своей индивидуальности на протяжении всего жизненного цикла клетки.
ДНК эукариот по структуре похожа на ДНК прокариот. Различия касаются:
- количества ДНК на геном;
- длины молекулы;
- порядка чередования нуклеотидных последовательностей;
- формы укладки ДНК. У прокариот – кольцевая форма молекулы ДНК, для эукариот - характерна линейная.
Особенностью ДНК эукариот является ее избыточность. У эукариот размер генов составляет около 1000 пар нуклеотидов. Генов у растений и животных много: у дрозофилы - 100 000, у млекопитающих и человека - 3-6 млн. нуклеотидных пар. А количество ДНК, участвующее в кодировании наследственной информации, необходимой для выполнения полной программы онтогенеза, - около 2% всей ДНК. Это является доказательством избыточности ДНК эукариот.
Часть избыточной ДНК представлена одинаковыми повторами нуклеотидов. Различают многократно и умеренно повторяющиеся последовательности нуклеотидов. Все они либо сконцентрированы в определенных местах генома и образуют структурный (конститутивный) хроматин, либо равномерно распределены вдоль генома. Структурный хроматин обычно встроен между уникальными последовательностями. Избыточная ДНК существует для управления генами, препятствует изменчивости.
Метафазная хромосома (спирализованный хроматин) состоит из двух хроматид. Форма определяется наличием первичной перетяжки - центромеры. Она разделяет хромосому на 2 плеча. Расположение центромеры определяет основные формы хромосом: метацентрические, субметацентрические, акроцентрические, телоцентрические.
Степень спирализации хромосом не одинакова. Участки хромосом со слабой спирализацией называют эухроматиновыми. Это зона высокой метаболической активности, где ДНК состоит из уникальных последовательностей. Зона с сильной спирализацией - гетерохроматиновый участок, способный к транскрипции. Различают конститутивный гетерохроматин-генетический инертный, не содержит генов, не переходит в эухроматин, а так же факультативный, который может переходить в активный эухроматин. Концевые отделы дистальных участков хромосом называют теломеры.
Хромосомы обладают следующими свойствами (правила хромосом):
1. Индивидуальности - отличия негомологичных хромосом.
2. Парности.
3. Постоянством числа - характерным для каждого вида.
4. Непрерывности - способности к репродукции.
Хромосомы подразделяются на аутосомы (соматических клеток) и гетерохромосомы (половых клеток).
По предложению Левитского (1924) диплоидный набор соматических хромосом клетки был назван кариотипом. Он характеризуется числом, формой, размерами хромосом. Для описания хромосом кариотипа по предложению С.Г. Навашина их располагают в виде идиограммы - систематизированного кариотипа. В 1960 году была предложена Денверская международная классификация хромосом, где хромосомы классифицированы по величине и расположению центромеры. В кариотипе соматической клетки человека различают 22 пары аутосом и пару половых хромосом. Набор хромосом в соматических клетках называют диплоидным, а в половых клетках - гаплоидным (он равен половине набора аутосом). В идиограмме кариотипа человека хромосомы делят на 7 групп, в зависимости от их размеров и формы.
1 - 1-3 крупные метацентрические.
2 - 4-5 крупные субметацентрические.
3 - 6-12 и Х-хромосома средние метацентрические.
4 - 13-15 средние акроцентрические.
5 - 16-18 относительно малые мета-субметацентрические.
6 - 19-20 малые метацентрические.
7 - 21-22 и Y-хромосома наиболее малые акроцентрические.
Согласно Парижской классификации хромосомы разделены на группы по их размерам и форме, а также линейной дифференцировке.
Хромосомы эукариот – это спирализованный хроматин – комплекс ДНК и белков, где 40% приходится на ДНК, 40% - на гистоновые (основные) белки и почти 20% - на негистоновые белки и немного РНК.
Гистоны - хромосомные белки с высоким содержанием аргинина и лизина. Гистоны стабилизируют структуру хромосомы и играют роль в регуляции активности генов.
Негистоновые (кислые) белки. В хромосомах их количество приблизительно вдвое меньше гистоновых. Существует более 100 видов негистоновых белков. Они разнообразны по молекулярному весу, структуре, видос-пецифичны. Эти белки могут быть ответственны за репликацию, репарацию, транскрипцию, возможно, играют роль и в активации генов. К ним относят актин, миозин, тубулин, ферменты синтеза РНК и ДНК - полимеразы и другие.
Из пяти классов гистонов, четыре (Н2А, Н2В, Н3, Н4) образуют своеобразные шаровидные тельца - нуклеосомы диаметром около 10 нм. В одну нуклеосому входит 8 молекул гистонов. Вокруг нуклеосомы уложен отрезок двуспиральной нити ДНК (около 140 нуклеотидных пар), который образует вокруг нее почти два оборота. Соседние нуклеосомы соединены друг с другом короткими отрезками ДНК (1-10 нм или 30-100 пар нуклеотидов). К каждому такому отрезку присоединены молекулы гистона Н1. На ген среднего размера приходится участок цепочки, приблизительно, из шести нуклеосом. Допускается, что вследствие взаимодействия Н1 с нуклеосомами происходит конденсация нуклеотидной структуры в особую хроматиновую нить (элементарную фибриллу, d=10 нм), которая закручивается в хроматиновую спираль (d= 25нм), или хроматиновый филамент. Хроматиновая спираль, возможно, идентична с хромонемой, которая во время деления скручивается в хроматиду.
Таким образом, уровни упаковки ДНК следующие:
1) Нуклеосомный (2,5 оборота двуспиральной ДНК вокруг восьми молекул гистоновых белков).
2) Супернуклеосомный - хроматиновая спираль (хромонема).
3) Хроматидный - спирализованная хромонема.
4) Хромосома - четвертая степень сперализации ДНК.
В интерфазном ядре хромосомы деконденсированы и представлены хроматином. Деспирализованный участок, содержащий гены, называется эухроматин (разрыхленный, волокнистый хроматин). Это необходимое условие для транскрипции. Во время покоя между делениями определенные участки хромосом и целые хромосомы остаются компактными.
Эти спирализованные, сильно окрашивающиеся участки, называются гетерохроматином. Они неактивны в отношении транскрипции. Различают факультативный и конститутивный гетерохроматин.
Факультативный гетерохроматин информативен, т.к. содержит гены и может переходить в эухроматин. Из двух гомологичных хромосом одна может гетерохроматической. Конститутивный гетерохроматин всегда гетерохроматичен, неиформативен (не содержит генов) и поэтому всегда неактивен в отношении транскрипции.
Хромосомная ДНК состоит из более 108 пар оснований, из которых образуется информативные блоки - гены, расположенные линейно. На их долю приходится до 25% ДНК. Ген - функциональная единица ДНК, содержащая информацию для синтеза полипептидов, или всех РНК. Между генами находятся спейсеры - неинформативные отрезки ДНК разной длины. Избыточные гены представлены большим числом - 104 идентичных копий. Примером являются гены для т-РНК, р-РНК, гистонов. В ДНК встречаются последовательности одних и тех же нуклеотидов. Они могут быть умеренно повторяющимися и высоко повторяющимися последовательностями. Умеренно повторяющиеся последовательности достигают 300 пар нуклеотидов с повторениями 102 - 104 и представляют чаще всего спейсеры, избыточные гены.
Высокоповторяющиеся последовательности (105 - 106) образуют конститутивный гетерохроматин. Около 75% всего хроматина не участвует в транскрипции, он приходится на высокоповторяющиеся последовательности и нетранскрибируемые спейсеры.
15. Ыидообразование
В территориальном аспекте в пространстве новый вид может возникнуть из одной или группы смежных популяций, расположенных на периферии ареала исходного вида. Такое видообразование называется аллопатрическим (от греч. alios — иной, patris — родина). В других случаях новый вид может возникнуть внутри ареала исходного вида, как бы внутри вида; этот путь видообразования называется симпатрическим (от греч. sym — вместе, patris — родина). В филогенетическом аспекте (во времени) новый вид может возникнуть посредством постепенного изменения одного и того же вида во времени, без какой-либо дивергенции исходных групп. Такое видообразование называется филетическим.
Новый вид может возникнуть путем разделения единого предкового вида (дивергентное видообразование). Наконец, новый вид может возникнуть в результате гибридизации двух уже существующих видов — гибридогенное видообразование. Кратко опишем эти основные пути видообразования.
Аллопатрическое видообразование (называемое иногда географическим) иллюстрируется приведенными выше примерами возникновения вида у больших чаек и в группе австралийских мухоловок. При аллопатрическом видообразовании новые виды могут возникнуть путем фрагментации, распадения ареала широко распространенного родительского вида. Примером такого процесса может быть возникновение видов ландыша (см. гл. 6). Другой способ аллопатрического видообразования — видообразование при расселении исходного вида, в процессе которого все более удаленные от центра расселения периферийные популяции и их группы, интенсивно преобразуясь в новых условиях, становятся родоначальниками видов. Примеры, аналогичные видообразованию в группе больших чаек, известны и для других птиц, некоторых рептилий, амфибий, насекомых.
В основе аллопатрического видообразования лежат те или иные формы пространственной изоляции, и этот путь видообразования всегда сравнительно медленный, происходящий на протяжении сотен тысяч поколений. Именно за такие длительные промежутки времени в изолированных частях населения вида вырабатываются те биологические особенности, которые приводят к репродуктивной самостоятельности даже при нарушении первичной изолирующей преграды. Аллопатрическое видообразование всегда связано с историей формирования видового ареала.
Симпатрическое видообразование. При видообразовании симпатрическим путем новый вид возникает внутри ареала исходного вида.
Первый способ симпатрического видообразования — возникновение новых видов при быстром изменении кариотипа, например при автополиплоидии. Известны группы близких видов (обычно растений) с кратными числами хромосом (см. рис. 6.28). Так, например, в роде хризантем (Chrysanthemum) все формы имеют число хромосом, кратное 9, 18, 27, 36, 45,..., 90. В родах табака (Nicotiana) и картофеля (Solanum) основное, исходное, число хромосом равно 12, но имеются формы с 24, 48, 72 хромосомами. В таких случаях можно предположить, что видообразование шло путем автополиплоидии — посредством удвоения, утроения, учетверения и т. д. основного набора хромосом предковых видов. Процессы полиплоидизации хорошо воспроизводятся в эксперименте посредством задержки расхождения хромосом в мейозе в результате воздействия, например, колхицином. Известно, что полиплоиды могут возникать и в природных условиях. Возникшие полиплоидные особи могут давать жизнеспособное потомство лишь при скрещивании с особями, несущими то же число хромосом (или при самоопылении). В течение немногих поколений в том случае, если полиплоидные формы успешно проходят «контроль» естественного отбора и оказываются лучше исходной диплоидной, они могут распространиться и сосуществовать совместно с породившим их видом (рис. 13.4) или, что бывает чаще, просто вытеснить его.
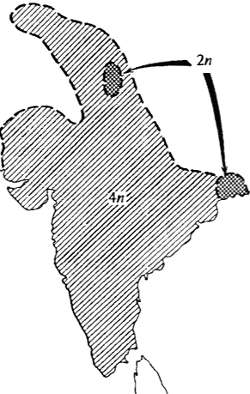
Рис. 13.4. Пример возникновения репродуктивной изоляции при полиплоидизации: обитающий по всему Индостану тетраплоидный вид растений Dicanthium annulatum, несомненно, возник из диплоидной предковой формы, ныне занимающей небольшой дизъюнктный ареал (по Н. Россу, 1962)
Полиплоидные формы, как правило, крупнее и способны существовать в более суровых физико-географических условиях. Именно поэтому в высокогорьях и в Арктике число полиплоидных видов растений резко увеличено (рис. 13.5). Среди животных полиплоидия при видообразовании играет несравненно меньшую роль, чем у растений, и во всех случаях связана с партеногенетическим способом размножения (например, у иглокожих, членистоногих, аннелид и других беспозвоночных).
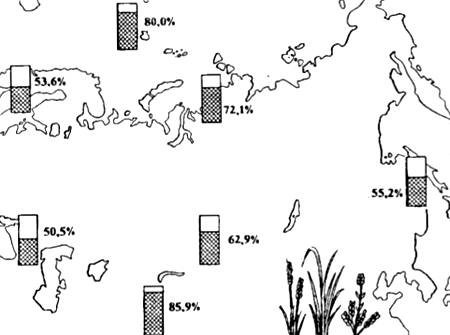
Рис. 13.5. Распространение полиплоидных видов цветковых (в процентах к общему числу видов флоры) в разных частях Евразии (по данным разных авторов из Н.В. Тимофеева-Ресовского и др., 1977)
Второй способ симпатрического видообразования — путем гибридизации с последующим удвоением числа хромосом — аллополиплоидия (см. ниже).
Наконец, последним, достаточно изученным способом симпатрического видообразования является возникновение новых форм в результате сезонной изоляции. Известно существование ярко выраженных сезонных рас у растений, например у погремка Alectorolophus major (см. гл. 10), раннецветущие и позднецветущие формы которого полностью репродуктивно изолированы друг от друга, и, если условие отбора сохраняется, лишь вопрос времени — когда эти формы приобретут ранг новых видов. Аналогично положение с яровыми и озимыми расами проходных рыб; возможно, что эти формы уже являются разными видами, очень схожими морфологически, но изолированными генетически (виды-двойники).
Особенность симпатрического пути видообразования — возникновение новых видов, морфофизиологически близких к исходному виду. Так, при полиплоидии увеличиваются размеры, но общий облик растений сохраняется, как правило, неизменным; при хромосомных перестройках наблюдается та же картина; при экологической (сезонной) изоляции возникающие формы также обычно оказываются морфологически слабо различимыми. Лишь в случае гибридогенного возникновения видов появляется новая видовая форма, отличная от каждого из родительских видов (но также имеющая признаки, характерные для двух исходных видов).
Филетическое видообразование. При филетическом видообразовании вид, весь в целом изменяясь в чреде поколений, превращается в новый вид, который можно выделить, сопоставляя морфологические характеристики этих групп.
Филетическое видообразование включает стазигенез — развитие вида во времени с постепенным изменением одной и той же экологической ниши, и анагенез — развитие вида с приобретением каких-то новых принципиальных приспособлений, позволяющих ему образовать совершенно новую, более широкую экологическую нишу. Примером стазигенеза может быть развитие основного ствола верхнеплиоценовых моллюсков рода Giraulus (см. рис. 6.5).
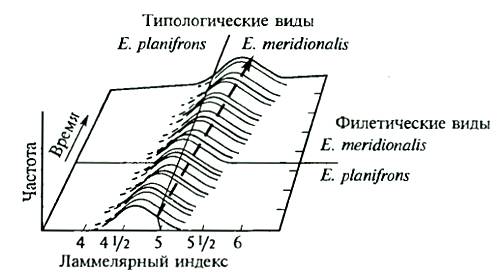
Рис. 13.6. Пример филетического видообразования в ряду ископаемых европейских слонов (Elephas planifrons — Е. meridionalis) по ламеллярному индексу (количеству эмали на зубах) (из В. Гранта, 1980)
Ясно, что в этом случае возможно лишь сопоставление морфологических характеристик, так как изучать результаты филетической эволюции можно лишь с привлечением палеонтологического материала (рис. 13.6). При этом всегда остается возможность того, что на каком-то этапе эволюции от единого филетического ствола могли дивергировать другие группы и филетическое видообразование могло оказаться на самом деле дивергентным. Поэтому в «чистом виде» филетическая эволюция, видимо, возможна лишь как идеализированное и упрощенное отражение эволюционного процесса на одном из отрезков жизни вида (фратрии).
Заметим, что границы между отдельными видами в филетическом ряду форм провести невозможно — она всегда будет условной (см. гл. 12).
Дивергентное видообразование (кладогенез). Ч. Дарвин считал этот тип видообразования самым распространенным (единственный рисунок в «Происхождении видов» посвящен именно этому типу видообразования). Примерами этого типа видообразования — возникновение новых видов в результате разделения единой предковой формы — являются возникновение нескольких видов сигов вокруг Ирландского моря (см. рис. 6.10) и дивергенция дарвиновых вьюрков на Галапагосах (см. рис. 6.12) и эволюция североамериканских дрозофил группы pseudoobscura — persimilis (см. рис. 6.27).
Гибридогенное видообразование (синтезогенез или сингенез). Этот тип видообразования обычен у растений: по некоторым подсчетам, более 50% видов растений представляют собой гибридогенные формы — аллополиплоиды. Укажем лишь на некоторые. Культурная слива (Prunus domestica) с 2n = 48 возникла путем гибридизации терна (P. spinosa, 2n = 32) с алычой (P. divaricata, 2n= 16) с последующим удвоением числа хромосом. Некоторые виды пикульника, малины, табака, брюквы, полыни, ириса и других растений — такие же аллополиплоиды гибридогенного происхождения.
Интересен случай возникновения нового симпатрического вида у Spartina townsendii (2л = 120) на основе гибридизации с последующим удвоением числа хромосом местного английского вида S. stricta (2n = 50) и завезенной в 70-х годах XIX в. из Северной Америки S. alternifolia (2n = 70). Сейчас ареал этого вида интенсивно расширяется за счет сокращения ареала местного европейского вида. Другой пример гибридогенного вида, возникшего симпатрически,— рябинокизильник (Sorbocotaneaster), сочетающий признаки рябины и кизильника и распространенный в середине 50-х годов в лесах южной Якутии по берегам среднего течения р. Алдан (К.М. Завадский). В результате гибридогенного видообразования особенно часто могут образовываться комплексы видов (или так называемых полувидов), связанных между собой гибридизацией,— сингамеоны (В. Грант). В случае таких гибридных комплексов иногда бывает трудно обнаружить четкие границы между отдельными видами, хотя виды как устойчивые генетические системы выделяются вполне определенно.
Все четыре основные формы видообразования во времени схематично показаны на рис. 13.7.
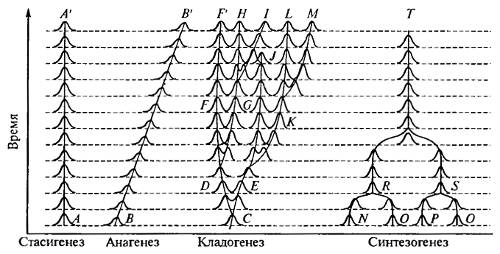
Рис. 13.7. Основные формы филетического (во времени) видообразования (из Н.Н. Воронцова, 2001)
Регуляции
Механизмы регуляции физиологических функций традиционно подразделяют на нервные и гуморальные, хотя в действительности они образуют единую регуляторную систему, обеспечивающую поддержание гомеостаза и приспособительную деятельность организма. Эти механизмы имеют многочисленные связи как на уровне функционирования нервных центров, так и при передаче сигнальной информации эффекторным структурам. Достаточно сказать, что при осуществлении простейшего рефлекса как элементарного механизма нервных регуляций передача сигнализации с одной клетки на другую осуществляется посредством гуморальных факторов - нейромедиаторов. Чувствительность сенсорных рецепторов к действию раздражителей и функциональное состояние нейронов изменяется под действием гормонов, нейромедиаторов, ряда других биологически активных веществ, а также простейших метаболитов и минеральных ионов (К+, Na+, Ca-+, С1~). В свою очередь, нервная система может запускать или выполнять коррекцию гуморальных регуляций. Гуморальные регуляции в организме находятся под контролем нервной системы.
Гуморальные механизмы филогенетически более древние, они имеются даже у одноклеточных животных и приобретают большое разнообразие у многоклеточных и особенно у человека.
Нервные механизмы регуляций образовались филогенетически и формируются постепенно в онтогенезе человека. Такие регуляции возможны лишь в многоклеточных структурах, имеющих нервные клетки, объединяющиеся в нервные цепи и составляющие рефлекторные дуги.
Гуморальные регуляции осуществляются путем распространения сигнальных молекул в жидкостях организма по принципу "всем, всем, всем", или принципу "радиосвязи".
Нервные регуляции осуществляются по принципу "письмо с адресом", или "телеграфной связи". Сигнализация передается от нервных центров к строго определенным структурам, например к точно определенным мышечным волокнам или их группам в конкретной мышце. Только в этом случае возможны целенаправленные, координированные движения человека.
Гуморальные регуляции, как правило, осуществляются медленнее, чем нервные. Скорость проведения сигнала (потенциала действия) в быстрых нервных волокнах достигает 120 м/с, в то время как скорость транспорта сигнальной молекулы с током крови в артериях приблизительно в 200 раз, а в капиллярах - в тысячи раз меньше.
Приход нервного импульса к органу-эффектору практически мгновенно вызывает физиологический эффект (например, сокращение скелетной мышцы). Реакция на многие гормональные сигналы более медленная. Например, проявление ответной реакции на действие гормонов щитовидной железы и коры надпочечников происходит через десятки минут и даже часы.
Гуморальные механизмы имеют преимущественное значение в регуляции процессов обмена веществ, скорости деления клеток, роста и специализации тканей, полового созревания, адаптации к изменению условий внешней среды.
Нервная система в здоровом организме оказывает влияние на все гуморальные регуляции, осуществляет их коррекцию. Вместе с тем у нервной системы имеются свои специфические функции. Она регулирует жизненные процессы, требующие быстрых реакций, обеспечивает восприятие сигналов, приходящих от сенсорных рецепторов органов чувств, кожи и внутренних органов. Регулирует тонус и сокращения скелетных мышц, которые обеспечивают поддержание позы и перемещение тела в пространстве. Нервная система обеспечивает проявление таких психических функций, как ощущение, эмоции, мотивации, память, мышление, сознание, регулирует поведенческие реакции, направленные на достижение полезного приспособительного результата.
17. Нервная регуляция
Нервная система объединяет, согласует и регулирует функции всех органов и систем, организует поведение человека, приспосабливая к меняющимся условиям среды, т.е. выполняет интегрирующую функцию. На любое раздражение организм реагирует как единая целостная система. Взаимодействие нейронов в ЦНС, обеспечивающее согласованную деятельность называется координацией. Она обусловлена рядом закономерностей – принципов координации, которые связаны как с особенностями строения, так и особенностями функционирования.
Морфологические принципы координации:
1. Принцип дивергенции заключается в том, что один нейрон, разветвляясь, контактирует посредством многочисленных терминалей с различными другими нейронами (рис. 3.9.1 Б). Такая особенность строения наиболее характерна для афферентных нейронов и лежит в основе процессов иррадиации.
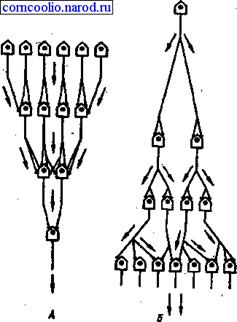
Рис. 3.9.1. А – конвергенция, Б – дивергенция.
2. Принцип конвергенции (принцип общего конечного пути, рис. 3.9.2) заключается в схождении аксонов от многих нейронов к одному эфферентному, который является для них общим (рис. 3.9.1 А). в ЦНС афферентных нейронов в 5 раз больше, чем эфферентных. Например, в головной мозг входят 2600000 афферентных волокон – выходят 140000 эфферентных.
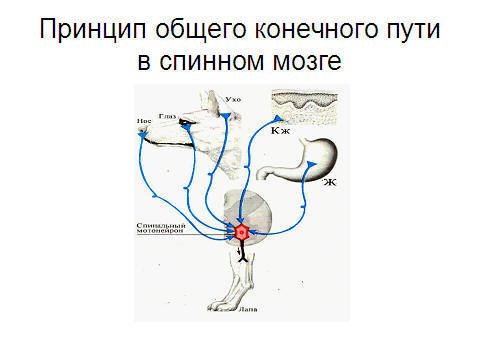
Рис. 3.9.2 К одному мотонейрону спинного мозга поступают нервные импульсы от различных рецепторов.
3. Наличие большого количества вставочных нейронов. Рефлекторные дуги и кольца имеют сложное строение за счет включения большого количества вставочных нейронов, которые располагаются как в спинном мозге, так и в различных отделах головного мозга. Этот принцип является морфологической основой иррадиации, обеспечивая большое количество вариантов ответной реакции, таким образом, обеспечивая адаптацию организма к изменяющимся условиям среды.
4. Наличие обратной связи. В ЦНС информация о выполнении рефлекторного акта, его соответствии условиям, вызывающим ответную реакцию, поступает по обратной связи (чувствительный нейрон, который несет импульс от рецепторов рабочего органа в нервный центр). Обратные связи представлены вторичными афферентными нервными волокнами, идущими в ЦНС от органов, по ним проходят нервные импульсы, возникающие в органах (тканях) во время их деятельности. ЦНС, получая импульсы, в любой момент может усиливать ответную реакцию (положительная обратная связь) или угнетает ее (отрицательная обратная связь). Обратная афферентация имеет большое значение в поддержании гомеостаза.
Физиологические принципы координации:
1. Иррадиация – при действии сильного и/или длительного раздражителя на определенные рецепторы, возбуждение (торможение) возникает не только в нервном центре данного рефлекса, но и распространяется от места возникновения на другие участки ЦНС (рис. 3.9.3 А, Б). Чем сильнее раздражение, тем больше выражен процесс иррадиации. В основе иррадиации лежит принцип дивергенции и наличие большого числа вставочных нейронов. Несмотря на широкие связи нервных центров, иррадиация имеет свои пределы и в деятельное состояние приходят лишь определенные отделы ЦНС.


Рис. 3.9.3. А – процесс возбуждения, Б – процесс торможения.
2. Концентрация заключается в постепенном сосредоточении нервного процесса к месту возникновения. В ее основе лежит принцип конвергенции. Концентрация протекает медленнее, чем иррадиация.

Рис. 3.9.4
3. Индукция – динамическое взаимодействие нервных процессов — возбуждения и торможения, выражающееся в том, что торможение в группе нервных клеток вызывает (индуцирует) возбуждение (положительная И.), и наоборот, первично вызванный процесс возбуждения индуцирует торможение (отрицательная И.).
4. Доминантные отношения. Поведение организма определяется жизненными потребностями. При усилении потребности возникает временно господствующий в ЦНС очаг возбуждения, нацеленный на удовлетворение именно этой потребности. Российский физиолог А.А. Ухтомский назвал такой механизм временного господства возбуждения доминантой. Доминанта является нейрофизиологической основой целенаправленного поведения (внимания, предметного мышления и т.п.) (рис. 3.9.5).

Рис. 3.9.5.
В естественных условиях доминантными становятся нервные центры, связанные с удовлетворением жизненно важных потребностей в данный период (пищевая, оборонительная, половая доминанты и т.п.).
Доминантный очаг обладает рядом свойств: 1) повышенная возбудимость, 2) способность привлекать на себя возбуждение из других отделов ЦНС, менее возбудимых в данный момент, 3) способность к суммации за счет притягиваемых импульсов, 4) оказывает сопряженное торможение на соседние центры, не входящие в очаг, 5) инерционность – стойкое и длительное сохранение возбуждения после прекращения действия раздражителя, вызвавшего доминанту.
Формирование условных рефлексов и их торможение связано также с возникновением в определенных отделах коры головного мозга доминантных очагов возбуждения. Доминанта имеет большое значение в педагогической деятельности.
Возрастные особенности координации. В первые дни жизни возбудимость понижена, затем она повышается и к концу 2-ой недели жизни становится больше, чем у взрослых. Ответная реакция у новорожденных и детей 1-го полугодия всегда сопровождается множеством ненужных движений, широкими неэкономичными вегетативными сдвигами. Незаконченная миелинизация нервных волокон обусловливает у детей высокую степень иррадиации. Кроме того, у детей в регуляции функций ведущую роль играют подкорковые отделы, так как не созрела кора больших полушарий. У детей легче и быстрее, чем у взрослых возникают, быстро сменяющиеся друг другом, доминантные очаги. У детей внимание неустойчивое, легко отвлекаются на самые различные раздражители. По мере функционального созревания ЦНС процессы координации у детей совершенствуются, полное созревание происходит к 18 – 20 годам.
Процессы саморегуляции основаны на использовании прямых и обратных связей. Прямая связь предусматривает выработку управляющих воздействий на основании информации об отклонении константы или действии возмущающих факторов. Например, раздражение холодным воздухом терморецепторов кожи приводит к увеличению процессов теплопродукции.
Обратные связи заключаются в том, что выходной, регулируемый сигнал о состоянии объекта управления (константы или функции) передается на вход системы. Различают положительные и отрицательные обратные связи. Положительная обратная связь усиливает управляющее воздействие, позволяет управлять значительными потоками энергии, потребляя незначительные энергетические ресурсы. Примером может служить увеличение скорости образования тромбина при появлении некоторого его количества на начальных этапах коагуляционного гемостаза.
Отрицательная обратная связь ослабляет управляющее воздействие, уменьшает влияние возмущающих факторов на работу управляющих объектов, способствует возвращению измененного показателя к стационарному уровню. Например, информация о степени натяжения сухожилия скелетной мышцы, поступающая в центр управления функций этой мышцы от рецепторов Гольджи, ослабляет степень возбуждения центра, чем предохраняет мышцу от развития избыточной силы сокращения. Отрицательные обратные связи повышают устойчивость биологической системы — способность возвращаться к первоначальному состоянию после прекращения возмущающего воздействия.
В организме обратные связи построены по принципу иерархии (подчиненности) и дублирования. Например, саморегуляция работы сердечной мышцы предусматривает наличие обратных связей от рецепторов самой сердечной мышцы, рецепторных полей магистральных сосудов, рецепторов, контролирующих уровень тканевого дыхания, и др.
Вся деятельность нервной системы осуществляется рефлекторным путем. С помощью рефлексов осуществляется взаимодействие различных систем целого организма и его приспособление к меняющимся условиям среды.
При повышении кровяного давления в аорте рефлекторно меняется деятельность сердца. В ответ на температурные воздействия внешней среды у человека суживаются или расширяются кровеносные сосуды кожи, под влиянием различных раздражителей рефлекторно меняется сердечная деятельность, интенсивность дыхания и т. д.
Благодаря рефлекторной деятельности организм быстро реагирует на различные воздействия внутренней и внешней среды.
Раздражения воспринимаются особыми нервными образованиями - рецепторами. Существуют различные рецепторы: одни из них раздражаются при изменении температуры окружающей среды, другие - при прикосновении, третьи - при болевом раздражении и т. п. Благодаря рецепторам центральная нервная система получает информацию обо всех изменениях окружающей среды, а также об изменениях внутри организма.
При раздражении рецептора в нем возникает нервный импульс, который распространяется по центростремительному нервному волокну и достигает центральной нервной системы. О характере раздражения центральная нервная система "узнает" по силе и частоте нервных импульсов. В центральной нервной системе происходит сложный процесс переработки поступивших нервных импульсов, и уже по центробежным нервным волокнам импульсы от центральной нервной системы направляются к исполнительному органу (эффектору).
Человек рождается с целым рядом готовых, врожденных рефлекторных реакций. Это безусловные рефлексы. К ним относятся акты глотания, сосания, чихания, жевания, слюноотделение, отделение желудочного сока, поддержание температуры тела и др. Количество врожденных безусловных рефлексов ограничено, и они не могут обеспечить приспособление организма к постоянно меняющимся условиям среды.
На базе врожденных безусловных реакций в процессе индивидуальной жизни формируются условные рефлексы. Эти рефлексы у высших животных и человека весьма многочисленны и играют огромную роль в приспособлении организмов к условиям существования. Условные рефлексы имеют сигнальное значение. Благодаря условным рефлексам организм заранее как бы предупреждается о приближении чего-то значимого. По запаху гари человек и животное узнают о приближающейся беде, пожаре; животные по запаху, звукам отыскивают добычу или, напротив, спасаются от нападения хищников. На основе многочисленных условных связей, образовавшихся в течение индивидуа








