| Доминантные признаки Рецессивные признаки
|
| Волосы: темные вьющиеся не рыжие
| Волосы: светлые прямые рыжие
|
| Глаза: карие большие
| Глаза:
голубые
маленькие
|
| Близорукость
| Нормальное зрение
|
| Ресницы длинные
| Ресницы короткие
|
| Нос с горбинкой
| Прямой нос
|
| Свободная мочка уха
| Приросшая мочка уха
|
| Широкая щель между резцами
| Узкая щель между резцами или ее отсутствие
|
| Полные губы
| Тонкие губы
|
| Наличие веснушек
| Отсутствие веснушек
|
| Шестипалость
| Нормальное строение конечностей
|
| Лучшее владение правой рукой
| Лучшее владение левой рукой
|
| Наличие пигмента
| Альбинизм
|
| Положительный резус-фактор
| Отрицательный резус-фактор
|
Применяемые Менделем приемы легли в основу нового метода изучения наследования — гибридологического.
Гибридологический анализ — это постановка системы скрещиваний, позволяющих выявить закономерности наследования признаков.
Условия проведения гибридологического анализа:
1) родительские особи должны быть одного вида и размножаться половым способом (иначе скрещивание просто невозможно);
2) родительские особи должны быть гомозиготными по изучаемым признакам;
3) родительские особи должны различаться по изучаемым признакам;
4) родительские особи скрещивают между собой один раз для получения гибридов первого поколения F1, которые затем скрещивают между собой для получения гибридов второго поколения F2;
5) необходимо проведение строгого учета числа особей первого и второго поколения, имеющих изучаемый признак.
234. Доминантные и рецессивные признаки.
Доминантный признак — признак, проявляющийся у гибридов первого поколения при скрещивании чистых линий. Результат наличия доминантного аллеля. Обычно «дикий тип», то есть вариант, присущий большинству особей природных популяций — это доминантный признак.
Рецессивный признак — признак, не проявляющийся у гетерозиготных особей вследствие подавления проявления рецессивного аллеля.
235. Аллельные гены.
Аллель – одна из форм одного и того же гена, определяет один из вариантов развития признака.
Обычно выделяют два аллеля: доминантный (соответствует нормальному гену) и рецессивный (объединяет в себя множество различных мутаций данного гена, приводящих к тому, что ген не работает).
При множественном аллелизме выделяют больше двух аллелей, например, в наследовании группы крови участвует три (IA, IB, I0). Диплоидный организм имеет два таких аллеля (один от папы, другой от мамы) в разных сочетаниях.
Организм, у которого аллельные гены одинаковы, называется гомозиготным (по данному признаку). Если аллельные гены разные – гетерозиготным. В гетерозиготе доминантный признак проявляется в фенотипе, а рецессивный – скрывается.
АЛЛЕЛЬНЫЕ ГЕНЫ (от греч. allélon — взаимно) — это гены, занимающие одинаковые локусы в гомологичных хромосомах и отвечающие за развитие альтернативных признаков. Аллельные гены, или аллели, представляют собой разные формы одного и того же гена. Обычно аллельных генов два: один — ген «дикого типа», доминантный и второй — его мутантная форма, рецессивный. Иногда мутантный аллель может доминировать над геном дикого типа (например, ген платиновой окраски меха у лис). Некоторые гены имеют несколько мутантных форм. В этом случае говорят о явлении множественного аллелизма. Серии множественных аллелей известны у дрозофилы (окраска глаз), у мышей (окраска шерсти), у кроликов (окраска меха) и др.
236. Фенотип и генотип.
Генотип – совокупность наследственных признаков и свойств, полученных особью от родителей. А также новых свойств, появившихся в результате мутаций генов, которых не было у родителей. Генотип складывается при взаимодействии двух геномов (яйцеклетки и сперматозоида) и представляет собой наследственную программу развития, являясь целостной системой, а не простой суммой отдельных генов. Целостность генотипа – результат эволюционного развития, в ходе которого все гены находились в тесном взаимодействии друг с другом и способствовали сохранению вида, действуя в пользу стабилизирующего отбора. Так, генотип человека определяет (детерминирует) рождение ребенка, у зайца – беляка потомство будет представлено зайчатами, из семян подсолнечника вырастет только подсолнечник.
Генотип – это не просто сумма генов. Возможность и форма проявления гена зависят от условий среды. В понятие среды входят не только условия, окружающие клетку, но и присутствие других генов. Гены взаимодействуют друг с другом и, оказавшись в одном генотипе, могут сильно влиять на проявление действия соседних генов.
Фенотип – совокупность всех признаков и свойств организма, сложившихся в процессе индивидуального развития генотипа. Сюда относятся не только внешние признаки (цвет кожи, волос, форма уха или нома, окраска цветков), но и внутренние: анатомические (строение тела и взаимное расположение органов), физиологические (форма и размеры клеток, строение тканей и органов), биохимические (структура белка, активность фермента, концентрация гормонов в крови). Каждая особь имеет свои особенности внешнего вида, внутреннего строения, характера обмена веществ, функционирования органов, т.е. свой фенотип, который сформировался в определенных условиях среды.
Если рассмотреть результаты самоопыления гибридов F2, можно обнаружить, что растения, выросшие из желтых семян, будучи внешне сходными, имеющие одинаковый фенотип, обладают различной комбинацией генов, т.е. разный генотип.
Понятия генотип и фенотип – очень важные в генетике. Фенотип формируется под влиянием генотипа и условий внешней среды.
Известно, что генотип отражается в фенотипе, а фенотип наиболее полно проявляется в определенных условиях среды. Таким образом, проявление генофонда породы (сорта) зависит от окружающей среды, т.е. условий содержания (климатические факторы, уход). Часто сорта, созданные в одних районах, мало пригодны к разведению в других.
237. Гомозигота и гетерозигота.
При изучении закономерностей наследования обычно скрещивают организмы, отличающиеся друг от друга альтернативными, т. е. взаимоисключающими признаками. Например, можно взять горох (как это сделал Мендель) с семенами желтыми и зелеными, морщинистыми и гладкими, окраской цветков пурпурной и белой, ростом растений высоким и низким.
Гены, от которых зависит развитие альтернативных признаков, принято называть аллельными, или аллелями (аллеломорфными парами). Аллельные гены расположены в гомологичных хромосомах в одних и тех же местах (локусах). Это, по сути, различные состояния одного и того же гена, определяющего внешние различия.

Аллельные гены. Гомозиготы и гетерозиготы.
Если в обеих гомологичных хромосомах находятся одинаковые аллели (например, оба кодируют желтую окраску семян или оба — морщинистую форму семян и т. д.), такой организм называется гомозиготным. Если же аллельные гены различны (например, в одной из гомологичных хромосом аллель кодирует желтый пигмент, а в другой гомологичной ей хромосоме аллель зеленого пигмента или один аллель— гладкой формы, а другой—морщинистой формы семян), то такой организм называется гетерозиготным.
Генотип и фенотип. Совокупность наследственных факторов организма (генов) называется генотипом. Совокупность всех признаков и свойств организма, являющихся результатом взаимодействия генотипа с условиями внешней среды, называется фенотипом. Поэтому организмы, имеющие одинаковый генотип, могут отличаться друг от друга в зависимости от условий развития и существования. Пределы, в которых в зависимости от условий среды изменяются фенотипические проявления генотипа, называются нормой реакции
238. Единообразие первого поколения.
Правило единообразия первого поколения: все гибриды первого поколения сходны между собой по признакам, полученным от родителей. В данном конкретном случае все растения F1 оказались с желтыми семенами, т. е. у них развился признак только одного родителя. Такие признаки, которые полностью преобладают в F1 называются доминантными. Противоположные им признаки, не проявляющиеся в F1 и развивающиеся лишь в определенной части особей в F2 и следующих поколениях, называются рецессивными. В рассматриваемом случае рецессивной оказалась зеленая окраска семян.
Правило единообразия подтверждается и для тех случаев, когда нет полного доминирования признаков одного родителя над признаками другого. Так, при скрещивании красноцветкового декоративного растения «ночная красавица» с белоцветковым все растения F1 оказываются розовыми (рис. 12), т. е. являются промежуточными по признаку окраски цветка между исходными формами. При этом правило единообразия первого поколения сохраняется.
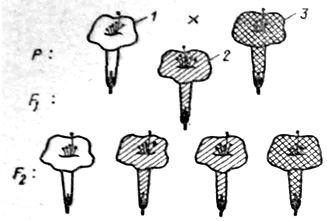
Рис. 12. Пример моногибридного скрещивания при неполном доминировании (скрещивание белой и красной ночных красавиц): 1 — белые цветки. 2 — розовые цветки, 3 — красные цветки
239. Промежуточный характер наследования.
Промежуточный характер наследования, или неполное доминирование, наблюдается в том случае, когда фенотип гетерозиготного гибрида отличается от фенотипа обеих родительских гомозиготных форм, т. е. выражение признака оказывается промежуточным, с большим или меньшим уклонением в сторону одного или другого родителя. Механизм этого явления состоит в том, что рецессивный аллель неактивен, а степень активности доминантного аллеля недостаточна для того, чтобы обеспечить нужный уровень проявления признака доминантной родительской гомозиготы.
Примером неполного доминирования является наследование окраски лепестков венчика у растений ночной красавицы. Гомозиготные растения имеют либо красные (АА), либо белые (аа) цветки, а гетерозиготные (Аа) — розовые. При скрещивании растения с красными цветками и растения с белыми цветками в F1 у всех растений цветки розовые, т. е. наблюдается промежуточный характер наследования. При скрещивании гибридов с розовой окраской цветков в F2 имеет место совпадение расщепления по фенотипу и генотипу, так как доминантная гомозигота (АА) отличается от гетерозиготы (Аа). Так, в разбираемом нами случае с растениями ночной красавицы расщепление в F2 по окраске цветков обычно следующее — 1 красная (АА): 2 розовые (Аа): 1 белая (аа).
Неполное доминирование оказалось широко распространенным явлением. Оно наблюдается в наследовании курчавости волос у человека, масти крупного рогатого скота, окраски оперения у кур, многих других морфологических и физиологических признаков у растений, животных и человека.
240. Закон расщепления признаков.
При скрещивании двух потомков первого поколения между собой (двух гетерозиготных особей) во втором поколении F2 наблюдается расщепление в определенном числовом соотношении: по фенотипу 3:1, по генотипу 1:2:1.
25% организмов, полученных во втором поколении F2, являются гомозиготными доминантными (АА), 50% — доминантны (Аа) по фенотипу и 25% — гомозиготны по рецессивному признаку (аа).
При неполном доминировании в потомстве гибридов F2 расщепление по фенотипу и генотипу составляет 1:2:1.
241. Статистический характер явлений расщепления.
Применение статистического метода в генетическом анализе позволило Менделю установить основные законы наследования.
Для того чтобы при скрещивании у животных и высших растений все фенотипические классы проявились в расщеплении, необходимо равновероятное образование разных сортов гамет и осуществление всех возможных их сочетаний при оплодотворении. Например, гибридное мужское растение Аа производит равное число пыльцевых зерен А и а, а женское, гомозиготное по а, производит одинаковые яйцеклетки. Однако при опылении на каждую яйцеклетку с аллелью а может приходиться как малое, так и большое число пыльцевых зерен. При малом их числе нет гарантии, что оба сорта пыльцы (А и а) окажутся в равном соотношении, так как одних пыльцевых зерен в силу случайности может быть больше, а других меньше. При большом же их числе вероятность того, что они будут с одинаковой частотой соединяться с яйцеклетками данного типа, возрастает.
Следовательно, важным условием реализации расщепления является размер, или объем, выборки, оцениваемой в опыте. Чем меньше количество особей в анализируемом потомстве, тем более вероятно случайное отклонение от нормального расщепления. Значение объема выборки еще более возрастает, если при оплодотворении сочетаются два сорта яйцеклеток — А и а и два сорта спермиев — А и а, так как здесь возрастает момент случайности. Чем больше элементов (сортов гамет) участвует в сочетании при прочих равных условиях, тем большего объема необходима выборка для наилучшего совпадения эмпирического расщепления с теоретически ожидаемым.
В гаплофазе расщепление учитывается на гаметах, а в диплофазе на зиготах и организмах. В последнем случае от мейоза до проявления признака протекают сложные процессы, которые могут маскировать результаты расщепления, при этом часть зигот может погибнуть; кроме того, признаки, определяемые генами, могут измениться под влиянием различных негенетических факторов.
Само явление расщепления является биологическим, а проявление его носит статистический характер. Поэтому в экспериментальной работе каждый раз необходимо убедиться в том, является ли обнаруженное отклонение от теоретически ожидаемого расщепления (1: 1; 3: 1; 9: 3: 3: 1) отклонением, вызванным закономерным влиянием каких-то факторов, нарушающих расщепление, или оно случайно, обусловлено, например, недостаточной величиной выборки.
В статистике принята считать, что если отклонение встречается чаще, чем 1 на 20 проб, то оно не случайно. Для оценки соответствия фактического расщепления теоретически ожидаемому необходимо определять значение отклонения.
Для статистической оценки отклонения применяют метод χ2 (хи-квадрат). Расчеты с помощью этого метода производят следующим образом. Сначала составляют таблицу по классам расщепления на основании опытных числовых данных. Затем из суммы частот всех классов, составляющей объем выборки, вычисляются теоретически ожидаемые величины (q) для каждого класса соответственно предполагаемой формуле расщепления (1:1, 3:1, 9: 3: 3: 1 и т. п.). Далее определяют отклонение (d) полученных данных от теоретически ожидаемых для каждого класса.
Каждое отклонение d возводят в квадрат, и квадрат отклонений (d2) делят на теоретически ожидаемое число (q) для данного класса:
d2/q
Все частные суммируют и получают величину χ2:
χ2 = Σ (d2/q)
Чтобы убедиться в том, что одинаковые отклонения могут иметь разное значение в зависимости от объема выборки, разберем пример расщепления по одной паре аллелей — черная и серая окраска мух при анализирующем скрещивании в опытах с дрозофилой. Теоретически ожидаемое отношение в данном скрещивании 1:1. Допустим, что один студент ограничивается в опыте 60 особями, другой получает больше — 300 особей. Как показывают данные, абсолютов отклонения одинаковы как для большой, так и для малой выборки, но величина χ2 гораздо больше для малой.
По методу χ2 можно определить вероятность того, насколько данное отклонение является случайным или, наоборот, закономерным. Производится это с помощью специальной таблицы Фишера.

Чтобы определить вероятность соответствия опытных и теоретически ожидаемых данных (Р) по данной величине χ2, необходимо сначала выяснить число степеней свободы. Отношения 1: 1 и 3: 1, т. е. в каждом по два класса, можно представить в виде суммы двух слагаемых, одно из которых устанавливается свободно, а второе — в зависимости от первого. В данном случае степень свободы может быть только одна. Чтобы было ясно, что такое степень свободы, приведем примеры.
В группе из 60 особей встречаются только два класса мух (черные и серые). Известно, что 40 из них серые. Если правильно отобрать 40 серых мух, то во втором классе автоматически будут только черные, и их должно быть 20, т. е. из суммы двух слагаемых одно взято свободно, а второе определилось автоматически. Если в F2 имеет место расщепление на три фенотипических класса (АА: Аа: аа), то после учета одного из них останется еще возможность выбора второго класса, а третий класс будет определен автоматически; из суммы трех слагаемых два берутся свободно, а третье имеет зависимое значение и т. д. Таким образом, число степеней свободы при анализе классов расщепления всегда будет на единицу меньше числа последних, т. е. если n — числа, классов, то число степеней свободы и будет равно n′ = n — 1.
В приведенном выше примере при двух классах расщепления степень свободы будет только одна. Вычислив величины χ2 отыскиваем соответствующие им вероятности для одной степени свободы по таблице Фишера. Так, для выборки в 60 особей, где χ2 = 6,6, вероятность Р будет меньше 0,01 т. е. соответствие теоретических и фактически полученных данных будет наблюдаться реже чем раз на 100. Однако, как мы уже говорили, в статистике считают явление случайным, если оно встречается реже, чем 1 раз на 20 проб (0,05). Иначе говоря, расщепление 40: 20 не соответствует 1:1. Во второй выборке в 300 особей χ2 = 1,34. Находя соответствующее ему значение вероятности по той же таблице, мы убеждаемся в совпадении опытных данных с теоретически ожидаемыми: 0,50 > Р > 0,20, т. е. Р = 0,2 ~ 0,5, что указывает на соответствие получаемых и ожидаемых частот примерно в 1 случае из каждых 2.
Тем же путем можно высчитать достоверность различий при дигибридном расщеплении по признакам семян гороха, полученном в опытах Менделя.
Метод χ2 дает возможность сравнивать различные численные отклонения при разных объемах выборок в одном масштабе, что очень важно для оценки опытных данных. Но следует иметь в виду, что этот метод неприменим, во-первых, к значениям, выраженным в процентах и относительных числах, и, во-вторых, к выборкам с числом особей в каком-либо из теоретически рассчитанных классов, меньшем, чем 5.
МСХА
242. Цитологические основы единообразия первого поколения и расщепления признаков во втором поколении.
243. Закон независимого наследования и его цитологические основы.
244. Закон сцепленного наследования Т. Моргана, его цитологические основы.
245. Полное и неполное сцепление.
Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
При неполном сцеплении гомологичные хромосомы могут обмениваться аллельными генами. Причиной этого является кроссинговер, который, в свою очередь, является результатом того, что при мейозе гомологичные хромосомы конъюгируют и могут обмениваться участками.
В результате этого при скрещивании дигетерозигот с генотипом  с гомозиготами по рецессиву, имеющими генотип
с гомозиготами по рецессиву, имеющими генотип  , в потомстве, наряду с обычными, появляется некоторое количество особей, образовавшихся в результате слияния кроссоверных гамет (рекомбинантов), имеющих генотип
, в потомстве, наряду с обычными, появляется некоторое количество особей, образовавшихся в результате слияния кроссоверных гамет (рекомбинантов), имеющих генотип  или
или 
246. Роль перекреста хромосом.
На кроссинговер влияет множество факторов как генетической природы, так и внешней среды. Поэтому в реальном эксперименте о частоте кроссинговера можно говорить, имея в виду все те условия, в которых она была определена. Кроссинговер практически отсутствует между гетероморфными Х - и Y -хромосомами. Если бы он происходил, то хромосомный механизм определения пола постоянно разрушался бы. Блокирование кроссинговера между этими хромосомами связано не только с различием в их величине (оно наблюдается не всегда), но и обусловлено Y -специфичными нуклеотидными последовательностями. Обязательное условие синапса хромосом (или их участков) — гомология нуклеотидных последовательностей.
Для абсолютного большинства высших эукариот характерна примерно одинаковая частота кроссинговера как у гомогаметного, так и гетерогаметного полов. Однако есть виды, у которых кроссинговер отсутствует у особей гетерогаметного пола, в то время как у особей гомогаметного пола он протекает нормально. Такая ситуация наблюдается у гетерогаметных самцов дрозофилы и самок шелкопряда. Существенно, что частота митотического кроссинговера у этих видов у самцов и самок практически одинакова, что указывает на различные элементы контроля отдельных этапов генетической рекомбинации в половых и соматических клетках. В гетерохроматических районах, в частности прицентромерных, частота кроссинговера снижена, и поэтому истинное расстояние между генами в этих участках может быть изменено.
Обнаружены гены, выполняющие роль запирателей кроссинговера, но есть также гены, повышающие его частоту. Они иногда могут индуцировать заметное число кроссоверов у самцов дрозофилы. В качестве запирателей кроссинговера могут выступать также хромосомные перестройки, в частности инверсии. Они нарушают нормальную конъюгацию хромосом в зиготене.
Обнаружено, что на частоту кроссинговера влияют возраст организма, а также экзогенные факторы: температура, радиация, концентрация солей, химические мутагены, лекарства, гормоны. При большинстве указанных воздействий частота кроссинговера повышается.
В целом кроссинговер представляет собой один из регулярных генетических процессов, контролируемых многими генами как непосредственно, так и через физиологическое состояние мейотических или митотических клеток. Частота различных типов рекомбинаций (мейотический, митотический кроссинговер и сестринские, хроматидные обмены) может служить мерой действия мутагенов, канцерогенов, антибиотиков и др.
247. Генотип как целостная исторически сложившаяся система.
Генотип — это совокупность генов данного организма. У человека около 100 тыс. генов.
Генотип как единая функциональная система организма сложился в процессе эволюции. Признаком системности генотипа является взаимодействие генов.
Аллельные гены (точнее, их продукты — белки) могут взаимодействовать друг с другом:
• в составе хромосом — примером является полное и неполное сцепление генов;
• в паре гомологичных хромосом — примерами являются полное и неполное доминирование, кодоминирование (независимое проявление аллельных генов).
Неаллельные гены взаимодействуют в следующих формах:
• кооперация — появление новообразований при скрещиваниях двух внешне одинаковых форм. Например, наследование формы гребня у кур определяется двумя генами:
R — розовидный гребень;
Р — гороховидный гребень.
Р RRpp х rrPP
розовидный гороховидный
F1 RrPp — появление ореховидного гребня в присутствии двух доминантных генов; при генотипе rrрр проявляется листовидный гребень;
• комплементарное взаимодействие — появление нового признака при наличии в генотипе двух доминантных не-аллельных генов. При таком взаимодействии во втором поколении возможно появление четырех вариантов расщепления. Примером является развитие антоциана (красящего пигмента) у цветков душистого горошка. При наличии в гомозиготном состоянии хотя бы одного рецессивного аллеля окраска не развивается и лепестки остаются белыми:

• эпистаз, или взаимодействие, при котором ген одной аллельной пары подавляет действие гена другой аллельной пары. Если в генотипе присутствуют два разных доминантных аллеля, то при эпистазе проявляется один из них. Проявившийся ген называют супрессором, подавляемый ген называют гипостатическим. При скрещивании двух белых кур (леггорн Ааbb и виандотт Aаbb) во втором поколении произойдет расщепление по фенотипу в отношении 13/16 белых — в тех случаях, когда в генотипе встречаются оба доминантных гена, или в случае полной рецессивности генотипа и 3/16 — окрашенных — в случаях, когда есть только один из доми-
нантных генов. В данном случае ген А подавляет ген В. В отсутствие гена А проявляется ген В и куры окрашены;
• полимерия — влияние на один признак нескольких не-аллельных однотипных генов. В результате степень выраженности признака зависит от числа доминантных аллелей разных генов в генотипе организма;
• плейотропия — влияние одного гена на развитие нескольких признаков. У человека известен ген, вызывающий появление паучьих пальцев (синдрома Марфана). Одновременно этот ген вызывает дефект в хрусталике глаза. Ген, вызывающий рыжую окраску волос, влияет на пигментацию кожи, появление веснушек.
248. Генетика пола.
П ол — это совокупность признаков и свойств организма, определяющих его участие в размножении.
Пол особи может определяться:
а) до оплодотворения яйцеклетки сперматозоидом (прогамное определение пола);
б) в момент оплодотворения (сингамное определение пола);
в) после оплодотворения (эпигамное определение пола).
До оплодотворения пол определяется у некоторых организмов в результате разделения яйцеклеток на быстро и медленно растущие. Первые (более крупные) после слияния с мужской гаметой дают самок, а вторые (мелкие) — самцов. У коловраток, способных размножаться помимо обычного полового размножения с оплодотворением, партеногенетически, часть партеногенетических яйцеклеток во время развития лишается половины хромосом. Из таких яиц развиваются самцы, а остальная часть дает начало самкам.
У морского кольчатого червя бонеллия определение пола происходит в процессе онтогенеза: если личинка садится на дно, из нее развивается самка, а если прикрепляется к хоботку взрослой самки, то самец.
У подавляющего же большинства эукариот пол закладывается в момент оплодотворения и определяется генотипически хромосомным набором, который зигота получает от родителей. Клетки мужских и женских особей животных организмов различаются по паре хромосом. Эту пару называют половыми хромосомами (гетеросомами) в противоположность остальным — аутосомам. Половые хромосомы принято обозначать как Х- и Y-хромо-сомы. В зависимости от их сочетания у  и
и  организмов различают 5 типов хромосомного определения пола:
организмов различают 5 типов хромосомного определения пола:
1) XX, ХО (O обозначает отсутствие хромосом) встречается у видов Protenor (насекомые);
2) XX, XY — он характерен, например, для дрозофилы, млекопитающих (в том числе и для человека);
3) XY, XX — этот тип определения пола характерен для бабочек, птиц, рептилий;
4) ХО, XX — наблюдается у тли;
5) гапло-диплоидный тип ( 2n, n) встречается, например, у пчел: самцы развиваются из неоплодотворенных гаплоидных яйцеклеток, самки — из оплодотворенных диплоидных.
Конкретные механизмы, связывающие развитие мужского или женского пола с определенным сочетанием половых хромосом у разных организмов различен. У человека, например, пол определяется наличием Y-xpo-мосомы: в ней есть ТДФ-ген, он кодирует тестикул - детерминирующий фактор, который определяет развитие мужского пола.
У дрозофилы же в Y-хромосоме находится ген фертильности, ответственный за плодовитость самца, а пол определяется баллансом числа Х-хромосом и числа наборов аутосом (обычный диплоидный организм содержит, соответственно, два набора аутосом). В Х-хромосомах расположены гены, определяющие развитие по пути самки, а в аутосомах — по пути самца.
Если отношение количества Х-хромосом к количеству наборов аутосом равно 0,5, то развивается самец, а если — 1, то самка.
Помимо нормальных самцов и самок иногда появляются интерсексы — особи, по своим половым признакам занимающие промежуточное положение между мужским и женским полом (не путать с гермафродитами!). Это может быть вызвано как анеуплоидией по половым хромосомам в гаметах, так и различными нарушениями (например, гормональными) в процессе дифференциров-ки пола.
У большинства организмов среди потомков получается 50 % самцов и 50 % самок, так как в норме расщепление по полу подчиняется законам обычного моногибридного скрещивания между гомо- и гетерозиготой, для гибридов которого характерно расщепление в соотношении 1:1.
Наследование и пол
В половых хромосомах, помимо генов, определяющих развитие пола, локализуются «обычные» фенотипические гены. Особенности их наследования определяются тем, что они составляют группу сцепления гетеросом. Явление сцепленного с полом наследования было открыто Т. X. Морганом, который обнаружил, что наследование окраски глаз у дрозофилы находится во взаимосвязи с полом родителей — результаты прямого и обратного скрещивания были неодинаковы. Проведя ряд экспериментов, ученый пришел к выводу, что в Y-xpo-мосоме самца не содержится участка, кодирующего окраску глаз.

У человека сцепленными с полом являются такие аномалии, как дальтонизм и гемофилия. Поскольку рецессивные гены этих заболеваний локализованы в Х-хромосомах, ими чаще болеют мужчины; женщины же обычно гетерозиготны и по этой причине не болеют.
Если ген локализован в Y-хромосоме, ему в клетке нет гомологичной аллели, такой организм называют гемизиготой.
Некоторые гены могут находиться и не в половых хромосомах, однако их проявление будет зависеть от пола особи: у одного пола признак проявится, у другого — нет. Такие признаки называют признаками, ограниченными полом. К ним относятся, например, наличие рогов у оленей (самцы рогаты, а самки безроги) или яйценоскость птиц, которая проявляется только у самок. Обычно проявление признака, ограниченного полом, зависит от гормонального статуса организма, в первую очередь, от соотношения половых гормонов.
249. Хромосомная теория наследственности.
Хромосомная теория наследственности
— сформулирована в 1911 г. американским ученым Т. Морганом. Ее сущность заключается в следующем:
— основным материальным носителем наследственности являются хромосомы с локализованными в них генами; - гены наследственно дискретны, относительно стабильны, но при этом могут мутировать;
— гены в хромосомах расположены линейно, каждый ген имеет определенное место (локус) в хромосоме;
— гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются совместно;
— число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
— сцепление генов может нарушаться в результате кроосинговера;
— частота кроссинговера прямо пропорциональна расстоянию между генами.
Значение этой теории заключается в том, что она дала объяснение законам Менделя, вскрыла цитологические основы наследования признаков и генетические основы теории естественного отбора.
250. Значение генетики для медицины и здравоохранения.
По данным мировой статистики, ежегодно среди новорожденных 4—5% детей имеют значительные наследственные дефекты. Всемирной организацией здравоохранения зарегистрировано свыше 1000 серьезных наследственных аномалий человека, связанных с нарушением жизненно-важных процессов.
В связи с повышением фона ионизирующей радиации и загрязнением окружающей среды мутагенами возрастает число вредных мутаций у человека.
Современный этап развития генетики открыл огромные перспективы направленного вмешательства в явления наследственности и селекции растительных и животных организмов, выявил важную роль генетики в медицине, в частности. в изучении закономерностей наследственных болезней и физических аномалий человека.
Задача современного человечества состоит в том, чтобы не только сохранить жизнь и здоровье ныне живущих людей, но и оградить будущие поколения от вредных мутаций.
Выяснение генетической природы отдельных наследственных заболеваний — важная задача настоящего и будущего медицины.
В 70-е годы XX столетия получили развитие целый ряд научных направлений, базирующихся на основных теоретических позициях генетики, селекции, а также смежных с ними наук: химия, физика, микробиология.
Возникли и успешно развиваются новые науки: биотехнология, генная инженерия, клеточная инженерия.
Использованием живых организмов и биологических процессов для промышленного производства разнообразных практически важных веществ занимается особая отрасль — биотехнология.
В последнее время в разработке биотехнологических процессов все чаше используют результаты исследований генной и клеточной инженерии.
Генная инженерия — прикладная отрасль молекулярной биологии, разрабатывающая методы перестройки геномов организмов с помощью выделения или введения отдельных генов или их групп.
Клеточная инженерия — отрасль биотехнологии, в которой применяются методы выделения клеток из организма, использование их в искусственных средах, а также гибридизация соматических клеток.
Перспективным направлением клеточной инженерии является клонирование (получение совокупности клеток или организмов от общего предка бесполым путем) организмов. Клонируют животных и некоторые растения.
Искусственное изменение организмов возможно в период их зародышевою развития. Этим занимается эмбриональная инженерия.
251. Вредное влияние никотина, алкоголя и наркотиков на наследственность человека.
Вся наследственная информация, которая передается от родителей детям и которая осуществляет выполнение генетической программы, двигающей по своим законам все процессы развития и функционирования живого организма, находится в молекулах ДНК (дезоксирибонуклеиновая кислота). Каждая молекула ДНК располагается в ядре клетки и является структурной единицей хромосом. В тоже время сама молекула ДНК состоит из нескольких десятков тысяч генов, а каждый ген является отдельной составной частью наследственного аппарата, т. е. несет в себе наследственную информацию о каком-либо конкретном индивидуальном признаке. По программе, заложенной в этих чудо-молекулах происходит формирование нашего организма – его рост и развитие как физическое, так и психическое (осознанное и неосознанное). Эта программа обеспечивает (направляет) движение всех жизненных процессов как внутри многочисленных видов клеток в нашем организме, так и организма в целом. Она наделяет нас индивидуальными особенностями, доставшимися нам по наследству от наших родителей. Один ген окрашивает наши глаза, другой — волосы. Программа, заложенная в ДНК, формир








