Независимо от способа получения гибридных семян по любой культуре сразу же в первом поколении необходимо определить по доминантным (маркерным) признакам отцовской формы действительность их гибридного происхождения, отбраковать возможные негибридные экземпляры, которые усложняют работу и снижают ее качество.
При работе с гибридным материалом необходимо всегда соблюдать правила изоляции особенно у перекрестноопыляющихся культур и факультативных перекрестников. С этой целью следует проводить индивидуальную изоляцию каждой семьи, а чаще всего для более правильной оценки гибридов у ржи, гречихи и других культур применяют метод резервов (половинок), при котором для оценки качества гибридов высеваются не все, а только часть гибридных семян, остальные храняться до следующего года в резерве. Семена лучших комбинаций этого резерва высеваются индивидуально или группами сходных комбинаций с соблюдением изоляции при дальнейшей селекционной работе. По первому поколению важно получить характеристику по показателю доминирования, проявлению степени гетерозиса. Эти данные позволяют судить о ценности и дельнейшей перспективе определенной комбинации.
С целью ускоренного размножения и получения наибольшего количества семян гибридов первого поколения их посев и уход нужно проводить самым тщательным образом на хорошо подготовленной и высоко плодородной почве с увеличенными площадями питания для растений.
Во втором поколении основная задача заключается во внимательном изучении проявления комбинационной изменчивости, отборе и всесторонней оценке выщепившихся форм, семена лучших из них должны составить новый исходный материал и высеваться в селекционном питомнике первого года (СП-1) по семьям для дальнейшей селекционной работы.
Таким образом, из вышеизложенного следует, что для использования метода внутривидовой гибридизации необходимы обширные знания селекционно-генетических вопросов и практические навыки по созданию гибридов запланированных комбинаций, умение правильно оценить полученные гибриды и использовать появляющиеся в процессе расщепления удачные комбинационные формы, новообразования и трансгрессии. Метод внутривидовой гибридизации имеет неисчерпаемые возможности по увеличению нового генофонда исходного материала. С его помощью создано огромное количество замечательных сортов на новом этапе синтетической селекции. Кроме вышеуказанных сортов, созданных методом гибридизации, можно еще назвать целый ряд замечательных сортов по каждой культуре для подтверждения огромной значимости метода внутривидовой гибридизации в современной селекции. Все 47 занесенных в Государственный реестр Беларуси сортов озимой и яровой пшеницы созданы методом отбора из различных гибридных комбинаций, 100% сортов озимой тритикале, овса, картофеля, люпина узколистного являются сортами гибридного происхождения. Селекция кукурузы, томатов, огурцов, сахарной и кормовой свеклы в основном базируется на создании гетерозисных гибридов F1.
Отдаленная гибридизация
Отдаленной гибридизацией называется такие скрещивания, когда подобранные пары принадлежат различным видам или родам, т.е. являются отдаленными не в географическом, а в родственном отношении. В соответствии с этим различают межвидовые (пшеница мягкая ´ пшеница твердая) и межродовые (пшеница ´ рожь) скрещивания. Отдаленной гибридизации принадлежит особая роль в эволюции и селекции. Под влиянием ее возникновения в естественных условиях и использования в практической деятельности процесс с выщеплением новых ранее не существовавших экземпляров, совмещающих признаки различных видов или родов за счет перекомбинаций наследственного материала и возникающих новообразований.
Первые опыты по отдаленной гибридизации растений были начаты в России в 1756 году И.Г. Кельрейтером. Он опубликовал результаты исследований в 1772 году по скрещиванию виргинского и перувианского видов табака. Полученные гибриды, удачно сочетали скороспелость, высокую урожайность и ценные качества обоих родителей. Из-за стерильности первого поколения созданные гибриды не нашли широкого применения, так как необходимо было ежегодно проводить такие скрещивания с целью получения гибридных семян для посева. С тех пор прошло уже более 250 лет, но интерес к получению новых растений с помощью отдаленной гибридизации, несмотря на то, что успехи от применения этого метода еще не многочисленные, не уменьшается, а значительно возрастает и приобретает все большие масштабы, усиливается его значимость.
Значение отдаленной гибридизации в создании нового исходного материала особенно велико на современном этапе селекционной работы, когда мы имеем выдающиеся успехи в деле создания весьма ценных сортов различных сельскохозяйственных культур.
Для дальнейшего прогресса в выведении новых сортов, обладающих комплексом биологических, хозяйственно-полезных и корреляционно связанных с ними морфологических признаков, очень часто возникает острая необходимость выйти за пределы вида для заимствования необходимых свойств от других видов. Например, создание иммунных сортов картофеля к фитофторозу, вирусным заболеваниям, раку, нематоде, колорадскому жуку за счет внутривидовой гибридизации практически невозможно, так как все многообразие сортов и форм культурного вида Solanum tuberosum не обладают устойчивостью к этим заболеваниям и вредителям. Но среди других видов рода картофеля такие формы имеются. К ним, прежде всего, относятся S.demissum, S.acaule, S.andigenum, S.vernei, S.maglea и другие.
За прошедший период, особенно в XX веке, по теории и практике отдаленной гибридизации накоплен богатый экспериментальный материал, выяснены неиспользованные возможности ее более широкого применения, определены объективные трудности осуществления этого типа скрещиваний, выявлены основные причины существующих генетических и других биологических барьеров, разработано ряд методов преодоления нескрещиваемости видов между собой и бесплодия первого поколения отдаленных гибридов.
Большой вклад в теорию и практику этого вопроса внесли такие ученые как И.В. Мичурин (методы преодоления нескрещиваемости плодово-ягодных культур), Г.Д. Карпеченко (плодовитые капустно-редечные гибриды), Н.В. Цицин (сорта пшенично-пырейных гибридов), А.Ф. Шулындин (первые сорта ржано-пшеничных амфидиплоидов) и другие.
При изучении исторического становления и эволюции (филогении) различных видов растений установлено, что в основе возникновения многих видов лежит естественная, спонтанная гибридизация между видами. Наиболее богатым и убедительным примером на этот счет может служить эволюция всего полиплоидного ряда пшеницы (рисунки 7 и 8) и создание человеком новой зерновой культуры тритикале с различными уровнями плоидности (рисунок 9). Эволюция видов пшениц группы эммеров шла по аналогичной схеме (рисунок 8). Тетраплоидные эммеры T. araraticum и T. timopheevi (AbAbGG) произошли из гибридов T. boeticum (AbAb) и Al. speltoides (GG) через полиплоидизацию и отбор при размножении.
Естественным гибридом и аллоплоидом от T. timopheevi и T. monocоccum является пшеница Жуковского (AbAbAbAbGG). Путем искусственной автополиплоидии T.timopheevi французский ученый Эсло получил октоплоидную пшеницу T.timonovum (AbAbAbAbGGGG). Грибобойная пшеница T. fungcidum (AuAuAbAbBBGG) является результатом скрещивания T.persicum и T.timopheevi, перевода гибридов на полиплоидную основу с отбором плодовитых потомств. При проведении отдаленной гибридизации селекционеру постоянно приходится сталкиваться с тремя основными проблемами: нескрещиваемость генетически отдаленных видов, невсхожесть гибридных семян и бесплодие полученных гибридов.

Рисунок 7 – Схема филогении рода Triticum L.
Эти проблемы возникают в связи с тем, что:
- пыльца не прорастает на рыльце другого вида;
- пыльца прорастает, но пыльцевые трубки растут медленно и не достигают зародышевого мешка;
- пыльцевые трубки достигают зародышевого мешка, но оплодотворения не происходит;
- оплодотворение происходит, но зародыш прекращает свое развитие на стадии нескольких клеток;
- зародыш при начальном нормальном развитии прекращает свое формирование, образуются невсхожие семена;
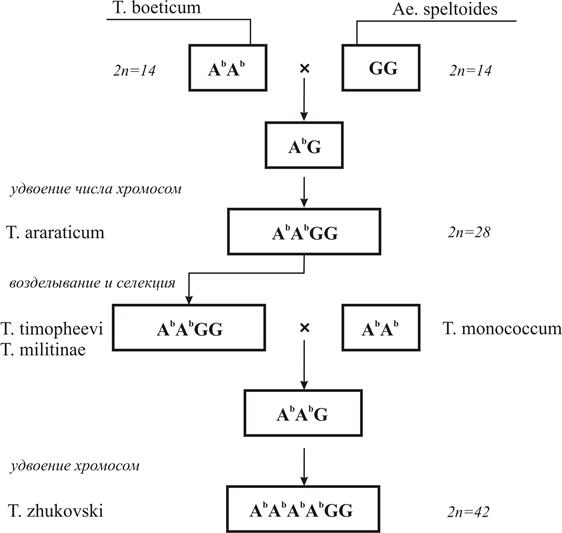
Рисунок 8 – Схема происхождения видов пшеницы группы эммеров
- при физиологическом несоответствии цитоплазмы и чужеродных хромосом, отсутствии парности хромосом скрещиваемых видов коньюгация не происходит, мейоз нарушается, половые клетки не образуются и гибридные
растения остаются бесплодными.
В деле преодоления нескрещиваемости подобранных пар видов для гибридизации или представителей различных родов в селекционной практике используются разработанные И.В. Мичуриным методы смеси пыльцы, предварительного вегетативного сближения, трансплантации частей столбика с рыльцем отцовского цветка, укорачивания столбика, метод посредника, реципрокных скрещиваний, нанесения биостимуляторов на рыльце пестика. В повышении эффективности отдаленной гибридизации большое значение имеет применение более новых методов, с помощью которых осуществляется предвари-
тельный перевод одного из компонентов на другой уровень плоидности с помощью полиплоидизации или деполиплоидизации. В результате этого оба родителя будут иметь одинаковое число хромосом и лучше скрещиваются
между собой. В тех случаях, когда оплодотворение происходит, но через несколько дней развитие зародыша прекращается, положительные результаты может дать метод эмбриокультуры, при которой в предкритический момент после оплодотворения семяпочка извлекается из завязи и трансплатируется на питательную среду in vitro. Этот метод целесообразно применять и в том случае, когда семена формируются полностью, но оказываются невсхожими.
Для преодоления бесплодия отдаленных гибридов первого поколения наиболее широко прибегают к возвратным скрещиваниям с одним из родителей или опылению пыльцой других видов. Успех обеспечивается в том случае, когда стерильность гибридов проявляется только по мужской части. В большинстве же случаев преодоление этой трудности осуществляется через удвоение числа хромосом гибрида первого поколения до фазы формирования генеративных органов. В этом случае восстанавливается парность гомологичных хромосом родительских видов, благодаря чему мейоз и весь процесс гаметогенеза у гибрида проходит более благополучно и делает его плодовитым. Плодовитые гибриды могут возникать и в том случае, когда при оплодотворении встречаются единичные нередуцированные гаметы. При этом образуются естественные плодовитые амфидиплоиды (аллополиплоиды). Заслуживает внимания и новый метод, разработанный в научно-практическом центре НАН Беларуси по земледелию И.А. Гордеем, когда для получения тритикале, например, используются ценные сорта тетраплоидной ржи и полиплоидизированные растения мягкой, твердой или других видов пшеницы. В этом случае в генотипе гибрида объединяются сразу полные геномы обоих родителей, позволяющие образовывать полноценные фертильные растения.
Для вегетативно–размножающихся растений бесплодие гибридов не имеет существенного значения, так как их размножение для получения урожая осуществляется несвязанными с половым процессом клубнями, луковицами, корневищами, отводками, черенками, другими органами и частями полученных гибридных растений.
Значительные успехи достигнуты по целому ряду сельскохозяйственных культур благодаря применению метода отдаленной гибридизации и отбора наиболее ценных новых форм из гибридного материала различных комбинаций скрещиваний.
Богата примерами в этом отношении селекция яровой пшеницы. Известный высокоустойчивый к гессенской и шведской мухам сорт яровой твердой пшеницы Харьковская 46 явился результатом отбора из сложного гибрида, полученного при скрещивании линии N 5129 (пшеница тургидная ´ пшеница двузернянка) с твердой пшеницей волжской экологической группы. В научно-исследовательском институте сельского хозяйства Юго-востока известными селекционерами А.П. Шехурдиным и В.Н. Мамонтовой были получены сорта Сарроза (Саратовская розовая) и Сарруба (Саратовская рубиновая) от скрещивания сортов Белотурка (твердая) ´ Полтавка (мягкая). Из комбинаций Лютесценс 91 ´ Сарроза получен сорт Альбидум 24, при скрещивании которого с Лютесценсом 55/11 были получены знаменитые сорта Саратовская 29, Саратовская 36, Саратовская 39. В создании последующих сортов Саратовская 38, Саратовская 42 и Саратовская 44 участвовал сорт Сарруба. Из комбинации Сарроза ´ Цезиум 111 выведен сорт Заволжская. В Краснодарском НИИСХ им. П.П. Лукьяненко из гибридов популяции твердой пшеницы Гордеиформе 27 ´ пшеницу двузернянку выделена линия Н-1122, при скрещивании которой с первым родителем получен ценный сорт Ракета. Сорт Новосибирская 7 является также результатом межвидовой гибридизации от скрещивания разновидности Ферругинеум (мягкая) ´ Гордеиформе (твердая). Он был получен методом отбора после скрещивания линий Лютесценс 1487 из этой комбинации с сортообразцом Лютесценс 956.
Успехов в создании первых в мире сортов озимой твердой пшеницы академику Ф.Г. Кириченко удалось добиться путем гибридизации мягкой пшеницы сорта Одесская 3 с различными формами твердой пшеницы. Так были получены первые районированные озимые сорта твердой пшеницы Мичуринка, Новомичуринка и Одесская янтарная, Кристалл 2, Парус, Коралл одесский. В Азербайджане сорт твердой пшеницы Кяхраба 10, получен путем сложной гибридизации пшеницы польской с пшенично-ржаным гибридом. Сорт Севиндж является результатом скрещивания тургидной пшеницы с твердой. При создании сорта мягкой пшеницы Гюргана использовались мягкая, твердая и шарозерная виды пшеницы. В создании канадского сорта Оттава использовалась пшеница двузернянка. От скрещивания образца двузернянки Вернал с мягкой пшеницей получен сорт твердой пшеницы Стеварт в США.
Широко известны достижения академика Н.В. Цицина по использованию метода межродовой гибридизации пшеницы с пыреем, который позволил получить сорта озимой пшеницы ППГ-1, ППГ-186, ППГ-599 и солрта яровой пшеницы ППГ-172, Восток, Грекум 114. Сорт Грекум 114 широко районирован в Казахстане, Алтайском крае и Бурятии урожайность которого при государственном испытании на Пржевальском сортоучастке достигала свыше 100 ц/га..
Исключительно убедительными примерами эволюционной деятельности человека является создание новой зерновой культуры тритикале благодаря освоению методики получения межродовых ржано-пшеничных гибридов. При скрещивании различных видов пшеницы с рожью через полиплоидизацию созданы плодовитые ржано-пшеничные плодовитые аллополиплоиды (амфидиплоиды). Большой вклад в этой области внесли В. Римпау, Г.К. Мейстер, А.И. Державин, В.Е. Писарев, В.Н. Лебедев, А. Киш, А.Ф. Шулындин, В.Е. Росенкова, С.И. Гриб и другие. Полученные А.Ф. Шулындиным трехвидовые тритикале (АА1ВВ1RR) объединяют в себе целое ядро ржи (14 хромосом), третью часть (14 хромосом) ядра мягкой пшеницы и половину (14 хромосом) ядра твердой пшеницы. В институте генетики и цитологии Национальной академии наук Беларуси Л.В. Хотылевой, Л.Ф. Ходорцовой Л.Н. Каминской проводятся исследования по созданию новых форм тритикале с использованием геномно замещенных линий пшеницы, у которых геном D замещен геномами различных видов эгилопса.
Лучшими первыми районированными сортами тритикале являются АД-206, Дар Белоруссии, Михась, Мара (озимые), Инесса, Лана (яровые).
Имеются примеры создания путем отдаленной гибридизации сортов многолетней ржи Державина и многолетней пшеницы сорта Первенец Саратовского государственного аграрного университета.
При скрещивании византийского овса с обыкновенным овсюгом в Калифорнии получен устойчивый к стеблевой и корончатой ржавчинам сорт овса Сиерра и скороспелые сорта Рапид и Мейза. Широко распространенный пластичный сорт Льговский 1026 создан методом отбора из комбинации византийского овса (Avena bysantina) x овес посевной (Avena sativa).
Г.В. Пустовойт с сотрудниками в своей работе при создании устойчивых сортов подсолнечника к заразихе, ржавчине, склеротинии, подсолнечной огневке интенсивно использовала отдаленную гибридизацию между H.tuberosum и H.annuus.
В результате гибридизации топинамбура с подсолнечником в Украине и на Северном Кавказе создан так называемый тописолнечник, обладающий признаками обоих родителей с проявлением высокой степени гетерозиса по урожайности клубней и зеленой массы. Благодаря вегетативному размножению этих гибридов гетерозис передается всем последующим поколениям размножаемых клонов.
Свыше 250 сортов картофеля при селекции на устойчивость к вирусам, нематоде, раку, фитофторе, колорадскому жуку, пониженным температурам создано благодаря использованию многих диких видов этого весьма полиморфного рода (2n = 24, 36, 48, 60, 72, 96). Известный среднеранний сорт Детскосельский создан из комбинации отдаленных скрещиваний Ранняя роза × (S.demissum × S.andigenum). Исходным материалом для создания иммунного сорта Имандра послужила комбинация отдаленного гибрида сорта Юбель × S.andigenum. Сорт Фитофтороустойчивый получен путем сложного скрещивания сорта Император × [(S.demissum × Гранат) × Гранат]. От скрещивания Приекуотского раннего × S.demissum создан сорт Арина. Из аналогичной комбинации Северная роза × S. demissum выделился ценный сеянец, послуживший родоначальником сорта Вятка. С участием вида картофеля S. andigenum в мире создано 780 сортов, а всего с потомствами от его сортов выведено более 2000 сортов с устойчивостью к различным болезням и вредителям, обладающие высоким содержанием крахмала, холодостойкостью и другими положительными качествами.
Широкое производственное значение и практическое использование в последние годы получили сорго-судановые гибриды, которые отличаются более высокой урожайностью по сравнению с сорго и суданской травой, содержат повышенное количество сахаров и белков.
Путем скрещивания сурепицы (2n = 20) с листовой капустой (2n = 18) искусственно синтезирован рапс (2n = 38), отличающийся высокой урожайностью, повышенным содержанием масла и лучшей зимостойкостью.
Всемирно известными являются работы И.В. Мичурина и его последователей С. Горшкова, П.Н. Яковлева, М.А. Лисавенко, С.Ф. Черненко, Э.П. Сюбаровой и многих других селекционеров по созданию ценных плодово-ягодных культур, которые применяли не только подбор географически отдаленных пар для внутривидовых скрещиваний, но и отдаленной гибридизации.
Интересные данные по межвидовой гибридизации томата имеются в Болгарии, Италии, Нидерландах, США. Наибольшая ценность в качестве доноров устойчивости к фитофторозу и бурой пятнистости имеют томат смородинолистный и томат волосистый соответственно.
Попытки получения гибридов между кукурузой и теосинте показали возможность создания нового типа кукурузного растения с повышенным содержанием белка и незаменимых аминокислот, которых этой культуре не хватает.
По зернобобовым культурам имеются всего лишь единичные успехи получения межвидовых гибридов. К ним относится получение сорта гороха Воронежский с участием красно-желтого гороха, сорта узколистного люпина Сидерат 892 из комбинации Lupinus angustifolius x Lupinus linifolius L. и новой формы белого люпина (L. alva Гaran) с синими цветками и серыми семенами в результате гибридизации L. albus ´ L. vavilovi Atab. в Белорусской государственной сельскохозяйственной академии.
В последнее время успешно решается проблема получения плодовитого гибрида между пшеницей и ячменем, в результате чего получен новый вид зернового растения под названием Tritordeum. Имеются примеры успешного применения отдаленной гибридизации в селекции табака, махорки и других культур.
Потенциальная возможность отдаленной гибридизации далеко еще не использована, предстоит раскрыть многие неизвестные механизмы взаимодействия цитоплазмы и ядра различных видов при их объединении в одну зачаточную клетку, предстоит более глубоко изучить физиологические и биохимические процессы, происходящие при прорастании пыльцы на рыльце чужеродного рыльца, продвижения пыльцевой трубки до семяпочки, слияния спермиев с центральным ядром и яйцеклеткой и всех процессов в постгамный период до полного формирования жизнеспособного семени.
Исключительно важную роль в дальнейшем совершенствовании и повышении эффективности отдаленной гибридизации должна сыграть биотехнология. С помощью ее методов культуры клеток и тканей возможна разработка способов извлечения из завязей оплодотворенных семяпочек и доращивания их на искусственном субстрате до получения гибридного растения, которое не может возникнуть обычным путем, так как во многих случаях через несколько дней после оплодотворения завязь с оплодотворенной семяпочкой отмирает. В последние годы доказана возможность получения отдаленных гибридов путем соматической гибридизации при слиянии клеток различных видов после их освобождения от клеточных оболочек с последующим получением каллуса, его дифференциацией до образования корней, листьев, стеблей и целых растений.
Экспериментальный мутагенез
Мутационные изменения постоянно происходят в природе и служат одной их основных предпосылок эволюции органического мира, так как они связаны с наследственной основой организмов и передаются следующим поколениям. Первый естественный мутант был обнаружен немецким аптекарем Шпрингером, который более 415 лет тому назад в 1590 году нашел измененную форму чистотела с сильно рассеченными листьями и лепестками цветков. У французского садовода в 1761 году появился экземпляр земляники с цельнокрайними листьями, который существует и до сих пор как отдельная разновидность. Подобные примеры были описаны в разное время и на других видах растений.
Ч. Дарвин в теории эволюции этот вид изменчивости назвал неопределенным, затрагивающим любые признаки и свойства живых организмов и передаваемый по наследству. В связи с этим он отнес его к числу основных факторов эволюции.
И.И. Герасимов в 1889 г. экспериментальным путем получил изменения кариотипа нитчатой водоросли спирагиры путем воздействия шоковых температур. Через 10 лет Коржинским была опубликована книга с многочисленными примерами возникновения и использования мутационных изменений в практической деятельности человека. Затем в 1901 году голландским ботаником Гюго де Фризом было дано теоретическое объяснение явлению мутаций и предложена мутационная теория.
Г.С. Филипповым и Г.А. Надсоном в 1925 году были получены экспериментальные мутанты дрожжей, Л.Н. Делоне и А.А. Сапегин воздействием излучений вызвали наследственные изменения у пшеницы, Стадлер в США провел аналогичные эксперименты на кукурузе. В.В. Сахаров и М.Е. Лобашев доказали возможность получения мутаций воздействием химических веществ.
Дальнейшее развитие и использование экспериментального мутагенеза было направлено не только на углубление теоретических исследований, но и на практическое применение этого метода в селекционном процессе. Особые заслуги в получении ценных штаммов для микробиологической промышленности принадлежат С.И. Алиханяну и А.И. Раппопорту в разработке и использовании химических супермутагенов в селекции растений.
В качестве физических мутагенов применяются рентгеновские лучи, гамма-излучения радиоактивных веществ. Наследственные изменения могут вызывать ультрафиолетовый свет, шоковые температуры. Из химических мутагенов наиболее широко применяют нитрозо-метил мочевину (НММ), нитрозо-этил мочевину (НЭМ), диметилсульфат (ДМС), этилен имин (ЭИ) и другие вещества, способные в десятых и сотых долях процентов раствора вызывать изменения наследственного материала.
В результате мутагенеза возникают генные, хромосомные и геномные мутации, связанные с потерей, удвоением, вставками, изменением порядка чередования нуклеотидов в молекулах ДНК; транслокацией, инверсией, дупликацией, нехватками отдельных участков хромосом; изменением количества хромосом в виде гаплоидии, анеуплоидии и полиплоидии (рисунок 10).

Рисунок 10 − Классификация мутационной изменчивости
При возникновении делеций происходит утрата участка молекулы ДНК или хромосомы с определенным количеством наследственной информации, что сказывается на качестве потомства. Во время дупликации происходит повторение отдельных участков хромосом или молекулы ДНК, а при транслокации изменяются положение определенного сегмента хромосом (сдвиг) или перемещение его из одной хромосомы в другую. Трансверсия – мутация в дуплексной ДНК, когда пурин заменяется на пиримидин или наоборот. При инверсии происходит поворот участка генетического материала на 180о, в результате которого считывание наследственной информации будет происходить в обратном порядке и сказываться на свойства синтезируемого белка.
Искусственные мутации можно вызвать при облучении физическими мутагенами сухих или набухших семян различными дозами радиации от 1 до 100 тыс. кр. Для каждого вида растений необходимо оптимальные дозы, подбирать экспериментальным путем, так как чувствительность обрабатываемых объектов является различной. Малые дозы облучения в большинстве случаев носят стимулирующий характер, а большие приводят к нежелательным и даже губительным результатам. Состояние обрабатываемых семян также имеет существенное значение. Набухшие семена оказываются в несколько раз чувствительнее по сравнению с сухими. Получение экспериментальных мутантов может осуществляться и во время роста растений на специально построенных гамма-полях или при использовании для питания растений радиоизотопов фосфора Р32, кобальта Со60 и других радиоактивных веществ. Мутации могут возникнуть при возделывании сельскохозяйственных культур на почвах загрязненных радионуклидами. Мутации в этих случаях, как правило могут возникать в процессе формирования генеративных органов, прохождении мейоза и всего процесса гаметогенеза.
Химические мутагены применяются в 0,01–0,10 %-ной концентрации для намачивания семян в течение нескольких часов с последующей промывкой их в проточной воде.
В первом и последующих поколениях мутантов необходимо проводить работу с особой тщательностью, так как проявление мутационной изменчивости имеет определенные особенности, в зависимости от рецессивного или доминантного состояния смутировавшихся генов.
В первом поколении (М1) истинные мутации в связи с их рецессивностью не проявляются, а возникающие фенотипические изменения чаще всего представляют собой морфозы и не всегда наследуются в дальнейшем. В связи с этим различающиеся по морфологическим признакам растения можно разделить на группы для их проверки при пересеве без применения индивидуального отбора. Более тщательная оценка, анализ характера проявления мутационной изменчивости и отбор растений с измененными признаками и свойствами проводится во втором поколении (М2) для индивидуальной проверки в потомстве М3 по типу селекционного питомника первого года (СП-1). Далее проводится работа по общепринятой схеме селекции совместно с другим исходным материалом различного происхождения. При работе с перекрестноопыляющимися культурами необходимо строго соблюдать изоляцию и отбор по методу половинок (резервов).
Отобранные и доведенные до константного состояния мутантные образцы не всегда могут дойти до конкурсного или государственного сортоиспытания, но их выбраковку следует проводить весьма осторожно, так как отдельные из них могут нести некоторые весьма ценные признаки по длине вегетационного периода, устойчивости к полеганию, болезням и вредителям, содержанию белка, жира и других полезных веществ. Носители таких наследственных изменений могут стать ценными донорами или источниками необходимых признаков, которые можно передать будущим сортам путем использования метода гибридизации.
С помощью физического и химического мутагенеза за несколько последних десятков лет достигнуты определенные успехи в создании нового исходного материала и сортов сельскохозяйственных культур.
В литературе сообщается, что в мире методом искусственного мутагенеза создано около 200 сортов различных культур. Примерами могут служить сорта ячменя ярового Минский и Факел, полученные в результате воздействия мутагенами на семена известного сорта Московский 121. В Швеции созданы сорта Паллас и Мари, сорт Диамант – в Чехии, Гамма 4 – в Японии, сорта озимого ячменя Пеннад и Лютер – в США, Ютта – в Германии.
Сорта овса Пенирад, Флорад, Флорад 500, Флорад 501 созданы в США, сорт кормового овса под названием Зеленый – в Краснодарском НИИСХ. В Швеции созданы сорта гороха Строл, ярового рапса Регина, горчицы Примекс.
В России и Украине районированы сорта мутантного происхождения яровой пшеницы Новосибирская 67, люпина Киевский мутант, гибрид кукурузы Краснодарская 82, Краснодарская 303 ВЛ, оливковый мутант подсолнечника Первенец, сои Универсал, картофеля Рентгеновский ранний, помидоров Луч 1, фасоли Сапарке 75, озимого ячменя Краснодарский мутант 1 (КМ-1) и сортообразец 33 М9. Из сорта ярового ячменя Краснодарский 35 методом мутагенеза получен скороспелый сорт Темп, из которого путем внутрисортового отбора создан устойчивый к полеганию короткостебельный сорт Каскад.
В Беларуси на Гомельской ОСХОС из мутантной популяции отобран сорт белого люпина Сож. В потомствах облученных семян сорта Трумпф выделен голозерный радиационный мутант ячменя Белорусский 76, отличающийся высокой урожайностью голого зерна со средним содержанием белка около 16 %. При дальнейшем облучении семян этого сортообразца лучами лазера получено ряд пленчатых линий, из которых в результате селекционной работы создан и доведен до районирования сорт Березинский. В экологическом и конкурсном испытаниях Белорусской государственной сельскохозяйственной академии он занял первое место по урожайности (8,2 т/га), превышение над стандартным сортом Роланд достигало 0,9 т/га.
В районировании находится литовский сорт льна-долгунца Балтучяй, выделенный из сорта Вайнгантас после обработки семян химическими мутагенами. Сорт сои Магева Рязанской ОСХОС получен методом мутагенеза.
Геномные мутации
К геномным мутациям относятся гаплоидия, анеуплоидия и полиплоидия, связанные с изменением числа хромосом в соматических и половых клетках.
Гаплоидный организм имеет одинарный (гаплоидный) набор хромосом (n). Такие организмы могут возникать в результате гаплоидного партеногенеза, когда яйцеклетка начинает развиваться без оплодотворения путем апомиксиса. Гаплоиды имеют только один геном, в котором каждый ген представлен в единственном числе, поэтому они представляют большой интерес с генетической точки зрения. Здесь все имеющиеся гены должны проявляться, ни один из них не может находиться в рецессиве, так как отсутствуют их доминантные аллели. Гаплоиды можно получить экспериментально путем стимуляции партеногенеза воздействием температурных и химических факторов. Хорошим примером получения гаплоидных растений является опыление цветков культурного ячменя пыльцой ячменя Hordeum bylbosum. После оплодотворения яйцеклетки зигота начинает развиваться, но геном ячменя бульбозум элиминируется в чужой цитоплазме и дальнейшее формирование зародыша происходит на гаплоидном уровне.
В настоящее время широкое применение для получения гаплоидных растений получил биотехнологический метод культуры пыльников, когда из пыльцевых зерен на искусственной среде удается получать растения с одинарным набором хромосом.
У гаплоидов при формировании половых клеток мейоз не происходит нормально, так как отсутствует парность гомологичных хромосом, без которых не может произойти коньюгация, а имеющиеся одиночные хромосомы ведут себя как униваленты. Вследствие этого для получения плодовитых растений гаплоиды необходимо переводить на диплоидную основу. Гаплоидные растения можно размножать только вегетативным путем клубнями, луковицами, черенками, отводками, усами, корневищами. Гаплоидия в селекции имеет большое значение, так с ее помощью можно полностью изучить генотип и сразу после перевода на диплоидный уровень создаются константные, гомозиготные по всем парам имеющихся генов исходные формы, называемые дигаплоидами.
Анеуплоидными называют растения, у которых имеется неправильный кариотип из-за добавления или нехватки целых хромосом. Среди анеуплоидов различают моносомики, в диплоидном наборе хромосом которых не хватает одной хромосомы (2n - 1); нулисомики характеризуются отсутствием пары гомологических хромосом (2n - 2) и полисомики, когда количество хромосом в соматических клетках на одну или несколько штук больше (2n + 1 – трисомики, 2n + 2 – тетрасомики, 2n + 1 + 1 – двойные трисомики).
Возникают анеуплоиды в результате нескольких дупликаций отдельных хромосом или при нарушении дупликации при митозе или мейозе. У всех анеуплоидов происходят нарушения в мейозе, поэтому они в большинстве случаев бывают стерильными.
Наибольший генетический и селекционный интерес представляют моносомики, которые создаются на основе любого сорта для проведения моносомного анализа с целью определения локализации определенных генов в хромосомах, находящихся в единичном состоянии. С этой целью создается серия моносомиков, количество которых равняется гаплоидному числу хромосом данного вида. У пшеницы, например, серия моносомиков состоит из 21 линии. Зная расположение необходимых генов в определенных хромосомах, для переноса нужных признаков при гибридизации используют моносомную линию по этой хромосоме. Моносомный анализ позволяет установить локализацию определенных генов в хромосомах и построить программу переноса и замещения нужных хромосом от сортов носителей ценных признаков внутри вида или за его пределы. Этим методом удалось сорту пшеницы Чайнз Спринг передать устойчивость к листовой ржавчине от эгилопса. Путем замены отдельных участков хромосом у мягкой пшеницы от пырея, эгилопсов и ржи удалось повысить ее устойчивость к стеблевой, бурой, желтой ржавчинам, твердой головне, мучнистой росе, полосатой мозаике.
Самыми значительными работами по созданию моносомных линий ряда сортов мягкой пшеницы являются работы Э. Сирса, О. И. Майстренко и других исследователей. К настоящему времени созданы полные ряды моносомиков (2n – 1) и трисомиков (2n +1) по многим культурам (таблица 1).
Таблица 1 − Ряды моносомиков и трисомиков у отдельных сельскохозяйственных культур.
| Культура
| Число хромосом
| Число хромосом в гаметах
|
| Полный ряд моносомиков (2n – 1)
|
| Пшеница – Triticum aestivum L.
| 42–1
| 21:20
|
| Овес – Avena sativa L.
| 42–1
| 21:20
|
| Табак – Nucotina tabacum L.
| 48–1
| 24:23
|
| Полный ряд трисомиков (2n +1)
|
| Пшеница – Triticum aestivum L.
| 42+1
| 21:22
|
| Овес – Avena sativa L.
| 42+1
| 21:22
|
| Рожь – Secale cereale L.
| 14+1
| 7:8
|
| Рис – Oruza sativa L.
| 24+1
| 12:3
|
| Сорго – Sordum vulgare L.
| 20+1
| 10:11
|
| Томат – Lycopersicom esculentym
| 24+1
| 12:13
|
| Шпинат – Spinacia oleracea L.
| 16+1
| 8:9
|
В результате кратного увеличения числа хромосом возникают полиплоидные организмы. Явление полиплоидии имеет важное исключительное эволюционое значение в растительном мире. Полиплоидные виды растений широко распространены в природе. С развитием цитологии было установлено существование определенного кариотипа для каждого вида растений. При изучении хромосомных наборов различных видов обнаружено, что в пределах определенного рода имеются полиплоидные серии или ряды видов с различной кратностью чис








