В соматосенсорную систему включают систему кожной чувствительности и чувствительную систему скелетно-мышечного аппарата, главная роль в которой принадлежит проприорецепции.
Кожа как сенсорная система. Кожа отграничивает организм от внешней среды; она содержит многообразные рецепторы, которые сигнализируют организму о контактах с внешним миром. Кожа состоит из трех слоев: эпидермиса, дермы (собственно кожи) и подкожной клетчатки. Под эпидермисом в слое соединительной ткани находится густая сеть нервных волокон с сенсорными окончаниями в эпидермисе и дерме. В дерме и подкожной клетчатке эти волокна собираются в пучки, а затем в крупные нервы, которые направляются в составе задних корешков в спинной мозг.
|
Рецепторные структуры кожи бывают двух видов: свободные и инкапсулированные окончания нервных волокон. Свободные окончания часто контактируют с клеткой-сателлитом эпидермального происхождения. Однако участвует ли эта клетка в реакции рецептора,- неизвестно. Самым простым типом рецептора кожи является свободный (не покрытый миелином) кончик тонкого афферентного волокна. Свободные нервные окончания находятся между клетками эпидермиса. По-видимому, значительная часть свободных окончаний представляет собой ноцицептивные рецепторы (болевые рецепторы). Другие свободные нервные окончания связаны с ощущениями зуда, щекотки; ими же, по-видимому, воспринимается температура (терморецепторы). Распространенным типом рецепторов являются также свободные нервные окончания, которые окружают волосяные фолликулы. Эти рецепторы возбуждаются при смещении волоса или его подергивании. В области волосяной фолликулы имеются также инкапсулированные окончания толстых миелинизированных волокон, которые, по-видимому, возбуждаются при сгибании волоса. Некоторые из немиелинизированных волокон являются эфферентными постганглионарными симпатическими волокнами, которые иннервируют гладкие мышцы сосудистой стенки и волосяные фолликулы (система обратной связи).
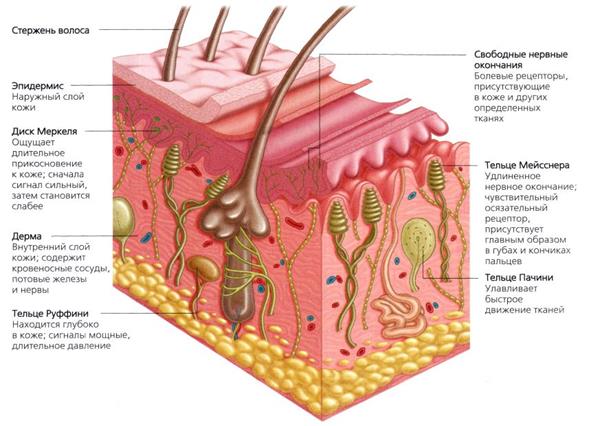
Рис. Рецепторный аппарат кожи
Инкапсулированные окончания составляют важный класс рецепторов кожи. Роль капсулы из соединительной ткани весьма значительна в формировании реакции рецептора. В настоящее время выделяют следующие инкапсулированные окончания кожи: диски Меркеля, тельца Мейснера, тельца Фатер-Паччини, окончания Руффини.
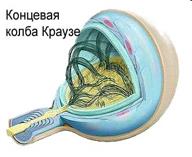
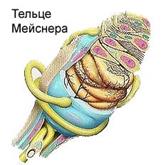

Рис. Примеры инкапсулированных рецепторов кожи.
При изучении свойств кожных рецепторов, ввиду большой плотности распределения и перекрытия их рефлекторных полей, трудно бывает сопоставить их физиологические характеристики с данными строения. Считается, что колбочки Краузе являются рецепторами холода, а тельца Руффини - рецепторы тепла; тельца Мейснера связаны с осязанием; Фатерпачиниево тельце чувствительно к давлению и вибрации, диски Меркеля – к прикосновению.
Рецепторы кожи можно разделить на быстро адаптирующиеся, или фазические (например, тельца Пачини); медленно адаптирующиеся, или тонические (например, диски Меркеля, окончания Руффини), и имеющие среднюю скорость адаптации, или фазно-тонические (например, рецепторы волосяных луковиц, тельца Мейснера). Большую роль в адаптации играет капсула рецептора: она ускоряет адаптационный процесс (укорачивает рецепторный потенциал), так как хорошо проводит быстрые и гасит медленные изменения.
Механизмы возбуждения кожных рецепторов. Механический стимул приводит к деформации мембраны рецептора. В результате этого электрическое сопротивление мембраны уменьшается, увеличивается ее проницаемость для Na+. Через мембрану рецептора начинает течь ионный ток, приводящий к генерации рецепторного потенциала. При увеличении рецепторного потенциала до критического уровня деполяризации в рецепторе генерируются импульсы, распространяющиеся по волокну в ЦНС.
Механорецепция объединяет четыре качества: ощущение прикосновения, давления, вибрации и щекотания. Ощущение прикосновения и давления на кожу довольно точно локализуется, т. е. относится человеком к определенному участку кожной поверхности. Эта локализация вырабатывается и закрепляется в онтогенезе при участии зрения и проприорецепции. Изучают механорецепцию одним из классических методов: прикосновением к коже волосками, или щетинками. Установлено, что даже легкое надавливание (в диапазоне 0,1-0,5 г, что приводит к вдавливанию кожи всего на 0,1-0,01 мм) вызывает ощущение прикосновения или давления. Но эти ощущения возникают только при надавливании в определенных точках, например кожи ладони. Эти точки называются тактильными точками. Участки кожи с большой плотностью тактильных точек находятся на кончиках пальцев и на губах. Плотность тактильных точек на животе, спине и плечах очень низкая.
Используя ножки специального циркуля (с тупыми концами, чтобы избежать болевого раздражения), можно определять пороги пространственного различения, т.е. минимальные расстояния между двумя одновременными тактильными стимулами, при которых их удается воспринять как раздельные. Исследования показали, что у взрослого человека одновременные пространственные пороги кончиков пальцев и губ особенно низки (1-3 мм), на слизистой оболочке языка порог пространственного различия равен 0,5 мм. На спине, плечах и бедрах они существенно выше (50-100 мм). Эти отличия обусловлены главным образом различными размерами кожных рецептивных полей (от 0,5 мм2 до 3 см2) и степенью их перекрытия.
Чувствительность к вибрации часто изучают при обследовании больных. Целью такого обследования чаще всего является нахождение порога субъективного ощущения вибрации. Наибольшая чувствительность к вибрации у человека находится в диапазоне 150 - 300 Гц. Минимальная (пороговая) амплитуда вибрации в этом частотном диапазоне составляет около 1 мкм.
Температурная рецепция. Температура тела человека колеблется в сравнительно узких пределах, поэтому информация о температуре окружающей среды, необходимая для деятельности механизмов терморегуляции, имеет особо важное значение. Терморецепторы располагаются в коже, роговице глаза, в слизистых оболочках, а также в ЦНС (в гипоталамусе). Они делятся на два вида: холодовые и тепловые (их намного меньше и в коже они лежат глубже, чем холодовые). Больше всего терморецепторов в коже лица и шеи. Терморецепторы можно разделить на специфические и неспецифические. Первые возбуждаются лишь температурными воздействиями, вторые отвечают и на механическое раздражение. Рецептивные поля большинства терморецепторов локальны. Терморецепторы реагируют на изменение температуры повышением частоты генерируемых импульсов, устойчиво длящимся все время действия стимула. Повышение частоты импульсации пропорционально изменению температуры, причем постоянная импульсация у тепловых рецепторов наблюдается в диапазоне температуры от 20 до 50°С, а у холодовых — от 10 до 41°С. Дифференциальная чувствительность терморецепторов велика: достаточно изменить температуру на 0,2 °С, чтобы вызвать длительные изменения их импульсации.
В некоторых условиях холодовые рецепторы могут быть возбуждены и теплом (выше 45°С). Этим объясняется возникновение острого ощущения холода при быстром погружении в горячую ванну. Важным фактором, определяющим установившуюся активность терморецепторов, связанных с ними центральных структур и ощущения человека, является абсолютное значение температуры. В то же время начальная интенсивность температурных ощущений зависит от разницы температуры кожи и температуры действующего раздражителя, его площади и места приложения. Так, если руку держали в воде температуры 27°С, то в первый момент при переносе руки в воду, нагретую до 25°С, она кажется холодной, однако уже через несколько секунд становится возможной истинная оценка абсолютной температуры воды.
Болевая рецепция. Болевая, или ноцицептивная, чувствительность имеет особое значение для выживания организма, так как сигнализирует об опасности при действии любых чрезмерно сильных и вредных агентов. В симптомокомплексе многих заболеваний боль является одним из первых, а иногда и единственным проявлением патологии и важным показателем для диагностики. Однако корреляция между степенью болевых ощущений и тяжестью патологического процесса отмечается не всегда.
Существуют специфические болевые рецепторы (свободные нервные окончания с высоким порогом реакции), кроме того, боль возникает при сверхсильном раздражении любых рецепторов. Болевые рецепторы в коже расположены в виде отдельных точек (дискретно). Поэтому в тех участках кожи, где их плотность невелика, всегда можно найти точки, лишенные болевых рецепторов. Специальные исследования показали, что кожа человека содержит значительно больше болевых точек, чем тактильных (соотношение 9:1).
Механизм возбуждения рецепторов при болевых воздействиях пока не выяснен. Предполагают, что особенно значимыми являются изменения рН ткани в области нервного окончания, так как этот фактор обладает болевым эффектом при встречающейся в реальных условиях концентрации ионов водорода. Таким образом, наиболее общей причиной возникновения боли можно считать изменение концентрации ионов водорода при токсическом воздействии на дыхательные ферменты или при механическом либо термическом повреждении клеточных мембран.
Не исключено также, что одной из причин длительной жгучей боли может быть выделение при повреждении клеток гистамина, протеолитических ферментов, воздействующих на глобулины межклеточной жидкости и приводящих к образованию ряда полипептидов (например, брадикинина), которые возбуждают окончания нервных волокон.
Адаптация болевых рецепторов возможна: ощущение укола от продолжающей оставаться в коже иглы быстро проходит. Однако в очень многих случаях болевые рецепторы не обнаруживают существенной адаптации, что делает страдания больного особенно длительными и мучительными и требует применения анальгетиков.
Болевые раздражения вызывают ряд рефлекторных соматических и вегетативных реакций. При умеренной выраженности эти реакции имеют приспособительное значение, но могут привести к тяжелым патологическим эффектам, например к шоку. Среди этих реакций отмечают повышение мышечного тонуса, частоты сердечных сокращений и дыхания, повышение давления, сужение зрачков, увеличение содержания глюкозы в крови и ряд других эффектов.
Понятие «боль» объединяет два типа соматических болей и висцеральные боли. Если соматическая боль имеет кожное происхождение, ее называют поверхностной болью. Если источник боли находится в мышцах, суставах или соединительной ткани, она называется глубокой болью. Таким образом, поверхностные и глубокие боли это два подкласса соматической боли.
Висцеральная боль (внутренних органов) также по своему характеру похожа на глубокую боль - имеет тупой диффузный характер. Примечательно, что внутренние органы (например, при вскрытии брюшной полости под местной анестезией) при сжимании и даже разрезах не вызывают ощущения боли, если только не задевать париетальную брюшину и корень брыжейки. Однако растяжение полых органов вызывает резкое ощущение боли. Кроме того, болью сопровождаются спазмы или сильные сокращения гладких мышц, особенно если этому сопутствует нарушение кровообращения (ишемия).
При ноцицептивных воздействиях на кожу человек локализует их достаточно точно, но при заболеваниях внутренних органов часты так называемые отраженные боли, проецирующиеся в определенные части кожной поверхности (зоны Захарьина—Геда). Так, при стенокардии, кроме болей в области сердца, ощущается боль в левой руке и лопатке.
Наблюдаются и обратные эффекты. Например, при локальных тактильных, температурных и болевых раздражениях определенных «активных» точек кожной поверхности включаются цепи рефлекторных реакций, опосредуемых центральной и автономной нервной системой. Они могут избирательно изменять кровоснабжение и трофику тех или иных органов и тканей. Поэтому оказываются эффективны методы иглоукалывания (акупунктуры), локальных прижиганий и тонического массажа активных точек кожи. Для уменьшения или снятия болевых ощущений в клинике используют множество специальных веществ — анальгетических, анестетических и наркотических. По локализации действия их делят на вещества местного и общего действия. Анестетические вещества местного действия (например, новокаин) блокируют возникновение и проведение болевых сигналов от рецепторов в спинной мозг или структуры ствола мозга. Анестетические вещества общего действия (например, эфир) снимают ощущение боли, блокируя передачу импульсов между нейронами коры большого мозга и ретикулярной формации мозга (погружают человека в наркотический сон).
В последние годы открыта высокая аналгезирующая активность так называемых нейропептидов, большинство из которых представляет собой либо гормоны (вазопрессин, окситоцин, АКТГ), либо их фрагменты (эндорфины). Аналгезирующее действие нейропептидов основано на том, что они даже в минимальных дозах (в микрограммах) меняют эффективность передачи в синапсах с «классическими» нейромедиаторами (ацетилхолин, норадреналин), в частности, между первым и вторым сенсорными нейронами (задние столбы спинного мозга и другие структуры).
Мышечная и суставная рецепция (проприорецепция). Мы можем достаточно точно оценивать положение конечностей, головы и туловища в пространстве. Кроме этого, достаточно точно оцениваются величина сопротивления каждому движению, углы суставов, скорость их изменения и пр. Такая оценка становится возможной благодаря рецепторам, которые находятся в мышцах, сухожилиях, суставах и называются проприорецепторами.
В мышцах млекопитающих животных и человека содержится три типа специализированных проприорецепторов: первичные окончания мышечных веретен, вторичные окончания мышечных веретен и сухожильные рецепторы Гольджи. Эти рецепторы реагируют на механические раздражения и участвуют в координации движений, являясь источником информации о состоянии двигательного аппарата организма.
Мышечное веретено представляет собой небольшое продолговатое образование длиной несколько миллиметров, шириной десятые доли миллиметра, расположенное в толще мышцы.
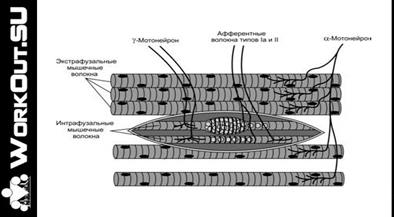
Рис. Мышечное веретено.
Каждое веретено покрыто капсулой. Внутри капсулы находится пучок мышечных волокон. Эти волокна называют интрафузальными в отличие от всех остальных волокон мышцы, которые носят название экстрафузальных. Веретена расположены параллельно экстрафузальным волокнам, поэтому при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении — уменьшается.
На интрафузальных волокнах спирально расположены чувствительные окончания афферентных волокон группы Iа — так называемые первичные окончания, и чувствительные окончания афферентных волокон группы II — так называемые вторичные окончания. Импульсация, идущая от веретен по афферентным волокнам группы Iа, в спинном мозге моносинаптически возбуждает мотонейроны своей мышцы и через тормозящий интернейрон тормозит мотонейроны мышцы-антагониста (реципрокное торможение). Афферентные волокна группы II возбуждают мотонейроны мышц-сгибателей и тормозят мотонейроны мышц-разгибателей. Имеются, однако, данные, что афферентные волокна группы II, идущие от мышц-разгибателей, могут возбуждать мотонейроны своей мышцы.
Веретена имеют и эфферентную иннервацию: интрафузальные мышечные волокна иннервируются аксонами, идущими к ним от γ-мотонейронов. Эти так называемые γ-эфферентные волокна подразделяют на динамические и статические. В расслабленной мышце импульсация, идущая от веретен, невелика. Веретена реагируют импульсацией на удлинение (растяжение) мышцы, причем у первичных окончаний частота импульсации зависит главным образом от скорости удлинения, а у вторичных — от длины мышцы (динамический и статический ответы). Активация γ-эфферентов приводит к повышению чувствительности веретен, причем динамические γ-эфференты преимущественно усиливают реакцию на скорость удлинения мышцы, а статические — на длину. Активация γ-эфферентов и без растяжения мышцы сама по себе вызывает импульсацию афферентов веретен вследствие сокращения интрафузальных мышечных волокон. Показано, что возбуждение α-мотонейронов сопровождается возбуждением γ-мотонейронов (α-γ-коактивация). Уровень возбуждения γ-системы тем выше, чем интенсивнее возбуждены α-мотонейроны данной мышцы, т. е. чем больше сила ее сокращения. Таким образом, веретена реагируют на два воздействия: периферическое — изменение длины мышцы, и центральное — изменение уровня активации γ-системы. Поэтому реакции веретен в условиях естественной деятельности мышц довольно сложны. При растяжении пассивной мышцы наблюдается активация рецепторов веретен, вызывающая рефлекс на растяжение. При активном сокращении мышцы уменьшение ее длины оказывает на рецепторы веретена дезактивирующее действие, а возбуждение γ-мотонейронов, сопутствующее возбуждению α-мотонейронов, вызывает активацию рецепторов. Вследствие этого импульсация от рецепторов веретен во время движения зависит от нескольких факторов: соотношения длины мышцы, скорости ее укорочения и силы сокращения.
Таким образом, веретена можно рассматривать как непосредственный источник информации о длине мышцы и ее изменениях, если только мышца не возбуждена. При активном состоянии мышцы необходимо учитывать влияние γ-системы. Во время активных движений γ-мотонейроны поддерживают импульсацию веретен укорачивающейся мышцы, что дает возможность рецепторам реагировать на неравномерности движения как увеличением, так и уменьшением частоты импульсации и участвовать таким образом в коррекции движений.
Сухожильные рецепторы Гольджи. Они находятся в зоне соединения мышечных волокон с сухожилием и расположены последовательно по отношению к мышечным волокнам. Сухожильные рецепторы слабо реагируют на растяжение мышцы, но возбуждаются при ее сокращении. Интенсивность их импульсации примерно пропорциональна силе сокращения мышцы, что дает основание рассматривать сухожильные рецепторы как источник информации о силе, развиваемой мышцей. Идущие от этих рецепторов афферентные волокна относятся к группе Ib. На спинальном уровне они через интернейроны вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антагониста.
Суставные рецепторы изучены меньше, чем мышечные. Известно, что суставные рецепторы реагируют на положение сустава и на изменения суставного угла, участвуя в системе обратных связей от двигательного аппарата и в управлении им.
Согласно современным данным, в суставном нерве выделяют два типа ответов: быстро адаптирующиеся и медленно адаптирующиеся, последние более многочисленны. Соответственно этому выделяют два типа рецепторов. Медленно адаптирующиеся рецепторы, являясь наиболее типичными суставными рецепторами, сообщают о положении костей, образующих данный сустав. В суставных нервах обнаруживаются волокна, в которых разряды возникают как при сгибании, так и при разгибании сустава. Обнаружены также волокна, разряд которых в зависимости от сгибания или разгибания сустава, учащается или урежается. Одно и то же движение сустава может вызвать урежение импульсации в одном афферентном волокне и увеличение ее в другом волокне.
Сигналы от рецепторов кожи, мышц, суставов, сухожилий и внутренних органов поступают через задние корешки в спинной мозг. Здесь эта информация обрабатывается и отсюда передается в верхние (супраспинальные) этажи ЦНС: ядра таламуса и далее в кору больших полушарий.
Сенсорные пути спинного мозга:
1. Лемнисковые пути задних столбов спинного мозга (тонкий и клиновидный пути) в эволюции формируются в связи с развитием конечностей, когда возникает необходимость в тонком анализе информации от суставов и кожи, без которого невозможны ходьба, бег, прыжки, удержание равновесия и положения тела в пространстве. Они тесно связаны с развитием соматосенсорной коры больших полушарий.
Путь передает в мозг сигналы о прикосновении к коже, давлении на нее и движениях в суставах. Отличительная особенность этого пути заключается в быстрой передаче в мозг наиболее точной информации, дифференцированной по силе и месту воздействия. Первые нейроны этого пути находятся в спинномозговом узле, их аксоны в составе задних столбов восходят к тонкому и клиновидному ядрам продолговатого мозга. В тонком ядре сосредоточены в основном вторые нейроны тактильной чувствительности, а в клиновидном ядре — вторые нейроны проприоцептивной чувствительности. Их аксоны перекрещиваются и, образуя медиальную петлю (лемниск), направляются в специфические ядра таламуса — вентробазальный ядерный комплекс. В этих ядрах концентрируются третьи нейроны лемнискового пути. Их аксоны направляются в соматосенсорную зону коры большого мозга.
По мере перехода на все более высокие уровни изменяются некоторые важные свойства нейронов лемнискового пути. Значительно увеличиваются (в продолговатом мозге в 2—30, а в коре большого мозга в 15—100 раз) размеры рецептивных полей нейронов. Отмечено появление так называемых нейронов новизны, реагирующих на смену раздражителя. Несмотря на увеличение размеров рецептивных полей, нейроны остаются достаточно специфичными (нейроны поверхностного прикосновения, глубокого прикосновения, нейроны движения в суставах и нейроны положения или угла сгибания суставов). Для корковой части лемнискового пути характерна четкая топографическая организация, т. е. проекция кожной поверхности осуществляется в кору большого мозга по принципу «точка в точку». При этом площадь коркового представительства той или иной части тела определяется ее функциональной значимостью: формируется хорошо известный сенсорный гомункулюс. Руки, особенно большой палец, оральная область, в том числе язык, голосовые связки представлены в коре мозга человека особенно подробно.
2. Спиноталамический тракт составляют самые тонкие миелинизированные афференты, а также немиелинизированные волокна задних корешков. Этот путь значительно отличается от лемнискового. Его первые нейроны также расположены в спинномозговом узле. Эти нейроны имеют большие рецептивные поля, иногда включающие значительную часть кожной поверхности. Вторые нейроны данного пути локализуются в сером веществе спинного мозга, а их аксоны в составе восходящего спинно-таламического пути направляются после перекреста на спинальном уровне в вентробазальный ядерный комплекс таламуса, а также в вентральные неспецифические ядра таламуса. Локализованные в этих ядрах третьи нейроны спинно-таламического пути лишь частично дают проекции в соматосенсорную зону коры. Спинно-таламический служит для передачи температурной, всей болевой и в значительной мере — тактильной чувствительности. В настоящее время считают, что этот тракт проводит генерализованные ощущения и по нему передается информация только о качестве раздражителя и его грубой локализации.
Болевая чувствительность практически не представлена на корковом уровне (раздражение коры большого мозга не вызывает боли), поэтому считают, что высшим центром болевой чувствительности является таламус, где 60 % нейронов в соответствующих ядрах четко реагирует на болевое раздражение.
Таким образом, в таламус проецируются две системы: задние столбы и спиноталамический тракт. Эти две системы различаются функционально: первая проводит информацию о месте и модальности раздражителя (протопатическая чувствительность), тогда как вторая обеспечивает генерализованные формы ощущения.
Нейрофизиология тройничного нерва. Ветви тройничного нерва осуществляют иннервацию различных тканей головы. Тройничный нерв (V пара черепно-мозговых нервов) - смешанного типа. Состоит из трёх ветвей:
1 - верхняя ветвь — глазничный нерв;
2 - средняя ветвь — верхнечелюстной нерв;
3 - нижняя ветвь — нижнечелюстной нерв.
Из них первые две чувствительные, третья содержит одновременно чувствительные и двигательные волокна.
Схема зон иннервации тройничного нерва
Зрительная ветвь иннервирует кожу лба и носа, роговицу, конъюнктиву и внутриглазные структуры, слизистую носовой полости, слезный проток. Верхнечелюстная ветвь иннервирует область кожи верхней челюсти, верхние зубы и десны, твердое небо и миндалины. Нижнечелюстная ветвь иннервирует область кожи нижней челюсти, слизистую щек, язык, проприорецепторы мышц нижней челюсти, нижние зубы и десны, жевательные мышцы. Все подразделения тройничного нерва иннервируют мозговые оболочки и сосуды мозга. Афферентные волокна тройничного нерва проводят афферентную импульсацию разной модальности - от механорецепторов, терморецепторов и болевых рецепторов. Окончания тройничного нерва образуют два типа терморецепторов: специфические терморецепторы реагируют только на изменение температуры, неспецифические - как на температурные, так и на механические воздействия. Пульпа зубов содержит только болевые рецепторы, которые иннервируются толстыми волокнами.
Чувствительные центростремительные волокна тройничного нерва представляют собой дендриты псевдоуниполярных нейронов мощного полулунного (гассерового) узла, который является гомологом спинномозговых ганглиев и содержит первые чувствительные нейроны тройничного нерва. Тела первых афферентных нейронов находятся также в среднемозговом тройничном ядре, которое представляет собой узкую полоску псевдоуниполярных нейронов, протянувшуюся вдоль всего среднего мозга. Это ядро, как и полулунном ганглий, является гомологом спинномозговых ганглиев. От них отходят аксоны к телам вторых нейронов, заложенных в ядрах в мозговом веществе ствола мозга. В зависимости от того, про какую чувствительность они несут информацию, переключение осуществляется в двух чувствительных ядра тройничного нерва.
Ядро болевой и тактильной (поверхностной) чувствительности представлено спинно-мозговым ядром тройничного нерва и является прямым продолжением задних рогов спинного мозга. Ядро имеет соматотопическую организацию.
Ядро проприоцептивной (глубокой) чувствительности – мостовое ядро тройничного нерва расположено в дорсолатеральной части покрышки моста.
Аксоны вторых нейронов идут в составе медиальной петли до таламуса. Переключение осуществляется в структурах как специфического, так и неспецифического отделов таламокортикальной системы (в заднемедиальном вентральном и интраламинарных ядрах таламуса). Ядра вентробазального таламуса переключают информацию о прикосновении, давлении, движении в суставах. Обработка этой информации на уровне специфических таламических ядер организована по принципу соматотопии и модальности. В отличие от этого для нейронов интраламинарных ядер, заднего и ретикулярного таламических ядер, к которым подходят также волокна спиноталамических трактов, характерна обширная конвергенция ряда модальностей, включая болевую.
Третьи нейроны тройничных путей, расположенные в таламусе, посылают свои аксоны через заднюю ножку внутренней капсулы к коре больших полушарий.
Соматосенсорная кора имеет два подразделения: SI (первичная соматосенсорная область) и SII (вторичная соматосенсорная область). В области SI коры выделяют есколько зон, соответствующих полям За, Зb, 1 и 2 (по Бродману). Нейроны поля За получают информацию от мышечных и суставных рецепторов, нейроны поля 2 - от глубоких структур (возбуждаются при сдавливании кожи и практически не реагируют на легкое прикосновение). В отличие от этого нейроны поля Зb реагируют на легкое прикосновение к коже. Нейроны поля 1 реагируют как на кожную стимуляцию, так и на стимуляцию глубоких структур. Характерным для сенсорных и моторных полей коры является организация нейронов, участвующих в обработке однородной информации, в вертикальные колонки. Например, нейроны одной колонки области SI реагируют на стимулы одной модальности. Кроме этого, соматосенсорные сигналы достигают нейронов моторной предцентральной коры (поля 4, 6) и поле 5 (ростральная теменная кора).
Эффекты нарушения сенсорных полей коры. Клинические наблюдения больных с поврежденной постцентральной областью коры показали, что у таких больных нарушается дискриминация размера и формы предметов. Например, больные не могли отличить на ощупь овал от параллелепипеда (в клинике это нарушение носит название астерогнозис). Как у человека, так и у обезьяны при повреждении постцентральной коры страдает кинестезия, что проявляется в резком ухудшении способности оценивать положение конечности при пассивном или активном ее перемещении. Страдает также оценка силы, приложенной к руке. Ощущение боли и температуры при этих повреждениях коры сохраняется.
При поражении вторичных отделов постцентральной коры у больных возникают своеобразные явления «афферентной апраксии». Они проявляются в том, что больной не способен выполнять тонкие дифференцированные движения. При поражении аналогичных зон левого (доминантного) полушария с распространением на нижние отделы (область представительства лица, губ и языка) кинестетическая апраксия может проявляться в нарушении движений речевого аппарата (афферентная моторная афазия). Нарушения проявляются в невозможности найти положение губ и языка, необходимые для артикуляции соответствующих звуков. Больные смешивают близкие артикулемы, например заменяют «б» или «п» на «м». Другими словами, больные смешивают акустически разные, но близкие по артикуляции буквы. При этих поражениях коры страдает также и письмо. Больной может вместо слова «халат» написать «хадат» или «ханат». Считают, что такие симптомы являются результатом нарушения афферентной основы движения.








