Отметим особенности дыхательного пигмента крови. Гемоглобин различных видов млекопитающих связывает одинаковое количество кислорода (1 г Hb c 1,34 см3 О2) [88]. Установлена также одинаковая концентрация гемоглобина в крови для различных видов млекопитающих (15 г на 100 мл крови) [214]. Показано [232], что форма кривых оксигенации гемоглобина у всех видов млекопитающих имеет S–образную форму. S-образность кривой имеет важное биологическое значение. Если бы кривые оксигенации имели, например, форму прямоугольной гиперболы, то при переходе к низким значениям рО2 происходила бы моментальная отдача кислорода в виде «взрыва». Кривые же S-образной формы обеспечивают диссоциацию оксигемоглобина не в форме «взрыва», а в виде постепенной и плавной отдачи кислорода на обменном участке (см. раздел 3.6, табл. 4). На молекулярном уровне S-образность формы означает, что связывание кислорода гемоглобином происходит кооперативно, т.е. присоединение кислорода к одному гему облегчает его присоединение к остальным.
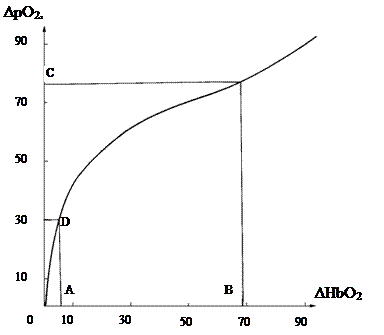
Кривая оксигенации гемоглобина HbO2=f(рО2), представленная на рис. 3.8, соответствует естественным параметрам крови: рН 7,40, рСО2=39,7 мм рт. ст. и t=37оС [176]. По мере продвижения эритроцита по цепочке обменных микрососудов непрерывно возрастает разность между значениями HbO2=97% и рО2=95 мм рт. ст. в аорте и их значениями в обменных микрососудах. На основе кривой HbO2= f(рО2) нами была построена разностная кривая DрО2=f(DHbO2) (см. рис. 4.3) [192], где DрО2 = 95-рО2 и DHbO2 = 97-HbO2. Разностной кривой соответствует система координат, в которой HbO2 и рО2 «поменялись» местами. При построении этой кривой за начало отсчета нами была принята точка с координатами HbO2=97% и рО2=95 мм рт. ст. на кривой HbO2=f(рО2). Кривая DрО2=f(DHbO2) отображает приращения DрО2 и DHbO2 в эритроците по мере прохождения последним цепочки обменных сосудов, включая и капилляр. Разностная кривая удобна для анализа закономерностей «кислородного разряда» эритроцита в капилляре. Произведенный нами анализ показал, что в пределах ΔHbO2 =5,5-68,6% и ΔpO2=30-76 мм рт. ст. (рис. 4.3, соответственно точки A, B, C, D) кривая ΔpO2=f(ΔHbО2) представляет собою сегмент логарифмической кривой [192]:
Δ pO 2= 42 lgΔ HbO 2. (4.21)
Всякой разности двух значений ΔHbO2 на логарифмическом участке кривой соответствует величина разности напряжения кислорода:
dpO 2= ΔрО 2(ΔHbO 2*) - ΔрО 2(ΔHbO 2^) = 42lg(ΔHbO 2*/ΔHbO 2^), (4.22)
где ΔHbO2*, ΔHbO2^ - значения ΔHbO2 на логарифмическом участке разностной кривой в пределах 5,5% £ ΔHbO2 £ 68,6%.
На кривой диссоциации гемоглобина в ее обычном представлении (см. рис. 3.8) логарифмическому» диапазону ΔpO2=30-76 мм рт. ст. соответствует интервал рО2=65-19 мм рт. ст., а ΔHbO2=5,5-68,6% интервал HbO2=91,5-28,4%. На артериальном конце капилляра в покое рО2 =50 мм рт. ст. [37, 38], а на венозном конце - рО2=20 мм рт. ст. [321]. Таким образом, в золотом режиме гипертензии диапазон рО2(bзс)=50-20 мм рт. ст. расположен в пределах «логарифмического» участка кривой диссоциации - 65-19 мм рт. ст. Напряжение кислорода на венозном конце капилляра совпадает с нижней границей логарифмического участка, а рО2 на артериальном конце ниже верхней границы логарифмического участка на 15 мм рт. ст. В условиях гипертензии рост среднего аортального давления в b раз вызывает уменьшение разности концентраций кислорода на участках аорта - артериальный конец капилляра и аорта - венозный конец капилляра в b раз. Эти изменения равнозначны уменьшению артерио-венозной разности насыщения
Рис. 4.3. Разностная кривая дезоксигенации гемоглобина человека [ 192 ] Размерность по оси абсцисс ΔHbO2 -. %, по оси координат ΔpO2 - мм рт. ст. Интервалы AB и CD соответствуют логарифмическому участку кривой, где ΔpO2=42lgΔHbO2. Интервал АВ - ΔHbO2=5,5-68,6%; интервал CD ΔpO2-=30-76 мм рт. ст. Кривая соответствует рН 7,40, рСО2=39,7 мм рт. ст. и температуре t =37оС.
на капилляре в b раз [192, 193]:
a- vHbO2 K(b) = a- vHbO2 K( b ЗС)/ b, (4.23)
где a-vHbO2K(bЗС), a-vHbO2K(b) - разность концентрации кислорода в крови на артериальном и венозном концах капилляра в золотом и других режимах гипертензии. С учетом (4.21)–(4.23), напряжение кислорода на концах капилляра при гипертензии возрастает на одну и ту же величину dpO2(b)=42lgb [192, 193]. Следовательно, напряжение кислорода на концах капилляра:
apO2(b) = 50 + 42 lgb, (4.24)
vpO2(b) = 20 + 42 lgb, (4.25)
где apO2(b), vpO2(b) - соответственно напряжение кислорода на артериальном и венозном концах капилляра в режимах гипертензии. В (4.24)-(4.25) отражены изменения рО2 в капилляре в различных режимах гипертензии. Максимальной гипертензии (b=2) соответствует приращение dpO2=12,6 мм рт. ст.; при этой гипертезии скорость эритроцитов в сердечном капилляре по отношению к золотому режиму возрастает в 2 раза. Интересно отметить, что при двукратном увеличении скорости эритроцитов в капиллярах аналогичное приращение (dpO2=10 мм рт. ст.) получено в различных точках нейрона мозга на модели Грюневальда [67]. Очевидно, что и при максимальном приращении величина рО2(b) не выходит за верхний предел логарифмического участка кривой насыщения гемоглобина (65 мм рт. ст.). Следовательно, величины рО2 на концах сердечного капилляра никогда не выходят за пределы логарифмического участка кривой. «Вхождение» в эти пределы обеспечивается изменениями в пределах гемодинамических параметров капилляра. Логарифмичность «капиллярного» участка кривой, как будет показано ниже, имеет большое значение для оптимального распределения величин рО2 в сердечной ткани. Исходя из (4.24) и (4.25), очевидно, что как в покое, так и при любой гипертензии артерио-венозная разность напряжений на капилляре:
a-vpO 2K ( b ) = 30 мм рт . ст. (4.26)
Благодаря большой крутизне «капиллярного» участка кривой эта разность минимальна, что обеспечивает наименьший разброс величин рО2 по объему призмы (см. раздел 4.5). Таким образом, при анализе кривой DрО2=f(DHbO2) нами установлена оптимальность «капиллярного» сегмента, на котором минимальному изменению DрО2 соответствует максимальное изменение DHbO2. На этом сегменте представлена оптимальная гармония «противоположностей» кислородных параметров по количеству и «качеству».
Необходимо отметить аналогию кислородного обеспечения по «количеству» и «качеству» в обменных артериолах и капиллярах. Каждое поперечное сечение капилляров и обменных микрососудов за единицу времени отдает одинаковое количество кислорода и в условиях покоя, и при гипертензии. Давление, приложенное к отдельным сосудам некоторой генерации обменных микрососудов одинаково, что обусловливает равное время пребывания эритроцитов в каждом из этих сосудов. Скорость отдачи кислорода из всякого поперечного сечения микрососудов постоянна и равна 220% с-1 (см. раздел 3.6). Наличие этих закономерностей означает, что рО2 на входе и выходе сосудов данной генерации приблизительно одинаковы. В конечном счете, феномен равенства обеспечивает одинаковую разность рО2 на входе и выходе всех капилляров. Величины рО2 на входе и выходе обменных сосудов различны и превышают таковые на капиллярах. Однако установлено [71], что уже на незначительном отдалении от стенки обменных артериол рО2 в ткани резко снижаются до приблизительно одинаковых величин. В то же время у стенок капилляров имеет место пологое падение рО2. На расстоянии свыше 10 мкм от стенки величины рО2 в ткани в обоих вариантах приблизительно одинаковы. Следовательно, «качество» и «количество» кислорода, поступающего в клетки из всех обменных микрососудов и капилляров при заданном уровне гипертензии, практически одинаковы для всех сосудов.
Таким образом, на примере тканевой призмы с ребрами-капиллярами нами проанализированы оба фактора кислородного обеспечения клеток: по «количеству» и «качеству». При увеличении среднего аортального давления в b раз «количество» кислорода обеспечивается за счет увеличения числа открытых микрососудов и капилляров в b раз, «качество» – за счет возрастания в b раз скорости эритроцитов в каждом капилляре [193]. Как следствие этих изменений, общий кровоток в капиллярной сети увеличивается в b2 раз.
Отметим, что обменные капилляры связаны между собою поперечными капиллярами; последние выполняют роль своеобразных анастомозов. Эту конструкцию можно рассматривать как своеобразный демпфер между открытыми капиллярами. Наличие этих устройств позволяет сбалансировать перепады в гемодинамике различных зон капиллярного русла. Установлено [84], что и в покое, и в условиях рабочей ишемии суммарная длина поперечных капилляров по отношению к суммарной длине всех постоянна и составляет 10%. Очевидно, что это постоянство не является случайным.
Отдает ли эритроцит на каждой единице длины капилляра одно и то же количество кислорода? Покажем, что такое постоянство существует. Отметим, что капилляр имеет форму усеченного кругового конуса (узкий диаметр на артериальном конце и широкий на венозном). Благодаря такой особенности в соответствии с принципом непрерывности эритроциты движутся в капиллярах с замедлением. Каждый эритроцит при движении отдает кислород с постоянной скоростью; однако время пребывания клетки в единичных сегментах капилляра различно. На узком единичном сегменте капилляра, который он успевает «проскочить» быстрее, эритроцит отдает кислорода меньше, чем на более широком. Однако за одно и то же время общее количество эритроцитов, «проскочивших» через узкий сегмент за единицу времени, больше, чем в широком. Вследствие этих обратных относительных изменений каждый единичный сегмент капилляра за единицу времени отдает приблизительно одинаковое количество кислорода.
Отметим еще одно важное обстоятельство. Очевидно, что в цилиндрическом капилляре эритроцит перемещался бы равномерно. В этом случае величина изменения насыщения гемоглобина на каждом единичном отрезке капилляра
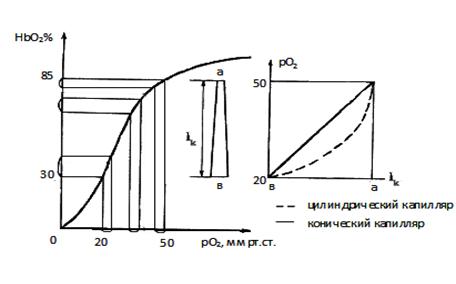
Рис. 4.4. «Распрямление» кривой дезоксигенации гемоглобина HbО2=f(pO2) на капилляре [ 194 ] . lk – длина капилляра, а, в артериальный и венозный концы капилляра.
(ΔHbO2/Δlк) была бы постоянной. При этом, исходя из формы кривой насыщения, величина ΔpO2/Δlк уменьшалась бы на каждой единице длины от артериального конца капилляра к венозному (см. рис. 4.4). Такое изменение ΔpO2/Δlк будет происходить при условии: капилляр - круговой цилиндр. Фактически же капилляр является круговым усеченным конусом, поскольку его диаметр на венозном конце больше, чем на артериальном, на 1-2 мкм [362]. Вследствие этого, отдельный эритроцит движется от начала капилляра к концу с возрастающим замедлением, отдавая на каждой единице длины все большее количество кислорода. Отметим, что при этом величина ΔpO2/Δlк меняется незначительно (см. рис. 4.4). Таким образом, отношение ΔpO2/Δlк остается приблизительно постоянным по всей длине капилляра. Нелинейный «логарифмический» сегмент кривой насыщения как бы «распрямляется» на капилляре. За счет этого феномена обеспечивается приблизительно линейное изменение величины рО2 по длине капилляров (см. рис. 4.4).
Таким образом, вследствие конусообразной формы капилляров и логарифмичности капиллярного участка кривой насыщения на единице длины капилляра обеспечивается: 1) диффузия одинакового объема кислорода за единицу времени, 2) линейное изменение величины рО2. Благодаря оптимальному сопряжению «противоположностей» кислородных параметров на каждом единичном сегменте капилляра создаются наиболее благоприятные условия для оптимального кислородного обеспечения призмы по «количеству» и «качеству».
Соотношение между внутренней поверхностью обменного микрососуда S и суммарной поверхностью SSЭ эритроцитов, проходящих мимо внутренней поверхности S, также имеет большое значение в явлениях газообмена. По расчетам, представленным А.Л. Чижевским [208], это соотношение убывает от крупных сосудов к капиллярам. Следует отметить, отношение SSЭ/S капиллярах имеет минимальное значение 1,26 (1,26@ 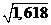 ). Нами показано (см. раздел 3.4.3), максимальный газообмен происходит в капиллярах, где имеет место наибольшее соотношение поверхности к объему сосуда (Sk/VK= max). Поэтому несомненно, что в капилляре при отношении SSЭ/SK=1,26 поверхность сосуда и cуммарная поверхность эритроцитов используются наиболее эффективно. Следует вспомнить, что в капиллярах эритроциты принимают парашютообразную форму [292], при которой эритроциты отдают большее количество кислорода, чем при дискообразной. Следовательно, наибольший уровень газообмена в капиллярах обусловлен не только оптимальным соотношением поверхности сосуда к его объему, но также и оптимальным соотношением поверхности сосуда к общей поверхности эритроцитов. Эти феномены составляет основу наиболее эффективного газообмена между капиллярами и сердечной тканью.
). Нами показано (см. раздел 3.4.3), максимальный газообмен происходит в капиллярах, где имеет место наибольшее соотношение поверхности к объему сосуда (Sk/VK= max). Поэтому несомненно, что в капилляре при отношении SSЭ/SK=1,26 поверхность сосуда и cуммарная поверхность эритроцитов используются наиболее эффективно. Следует вспомнить, что в капиллярах эритроциты принимают парашютообразную форму [292], при которой эритроциты отдают большее количество кислорода, чем при дискообразной. Следовательно, наибольший уровень газообмена в капиллярах обусловлен не только оптимальным соотношением поверхности сосуда к его объему, но также и оптимальным соотношением поверхности сосуда к общей поверхности эритроцитов. Эти феномены составляет основу наиболее эффективного газообмена между капиллярами и сердечной тканью.
4.5. Распределение рО2 в капиллярно-тканевыx призмах
В режимах гипертензии
Важнейшим фактором оптимизации кислородного обеспечения миокарда по «качеству» является и смещение концов соседних капилляров относительно друг друга. Представим взаимное расположение капилляров в виде прямой призмы, построенной на квадратном основании, с четырьмя перфузируемыми капиллярами в качестве ребер (мышечное волокно проходит внутри призмы) (см. рис. 4.1 и 4.2). Установлено [309], в миокарде кровотоки в соседних капиллярах в преобладающей степени противоположны. Отметим, потребление кислорода сердечной мышцей в течение систолы и диастолы резко различается [294]. Однако существует полная аналогия по «качеству» кислорода между «систолической» и «диастолической» призмами. Это подобие очевидно из следующих экспериментальных данных. В течение кардиоцикла распределение величин рО2 в крови сердечных капилляров [315], в отдельном кардиомиоците [353] и сердечной ткани [238] неизменно. Отметим, что величина среднего напряжения кислорода рО2 во всех регионах сердца в течение кардиоцикла одинакова [264, 269]. Отметим также, скорость отдачи кислорода из каждого сечения капилляров, открытых в тот или иной момент сердечного цикла, одинакова. Неизменность «качества» О2 в различных регионах в любую фазу кардиоцикла возможна только при одном условии. Этот феномен может иметь место, если существует равенство:
V D( b ЗС)/ V S( b ЗС) = B S( b ЗС)/ B D( b ЗС),
где VS(bЗС), VD(bЗС) – объем систолической и диастолической призм, ВS(bЗС), ВD(bЗС) – средняя скорость потребления кислорода единичным объемом «систолической» и «диастолической» призм за единицу времени в золотом режиме гипертензии. Поскольку призмы имеют квадратное основание и одинаковую высоту, то отношение расстояний между соседними капиллярами в «диастолической» и «систолической» призмах обратно пропорционально отношению скоростей потребления кислорода за систолу и диастолу:
đ D( b ЗС)/đ S( b ЗС) = 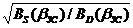 , (4.27)
, (4.27)
где đS(bЗС), đD(bЗС) – «систолическое» и «диастолическое» расстояния между капиллярами в золотом режиме гипертензии. В покое объем потребляемого кислорода миокардом за систолу составляет 91% и 9% в течение диастолы [294], а длительность диастолы превышает интервал систолы в 1,62 раз. Из этих данных следует, что «диастолическая» призма за единицу времени потребляет кислорода в 16,35 раз меньше, чем систолическая. Соответственно, по нашим расчетам отношение ВS(bЗС)/ВD(bЗС)=16,5, а đD(bЗС)/đS(bЗС)=4,07. При этих значениях обеспечивается одинаковое «качество» О2 в систолической и объемных призмах в любой момент кардиоцикла. Очевидно, что произведение объема систолической призмы на скорость потребления кислорода единичным объемом призмы равно аналогичному «диастолическому» произведению. Это произведение является своего рода инвариантом кислородного обеспечения в любой момент сердечного цикла. Этот инвариант является гарантом того, что по всему объему сердечной мышцы во всякий момент кардиоцикла обеспечивается адекватное кислородное обеспечение по «количеству» и «качеству».
Как было показано нами в разделе 4.4, конусообразность формы капилляра и логарифмичность «капиллярного» участка кривой насыщения обеспечивают одинаковое количество и «качество» потребляемого кислорода по всей длине открытого капилляра. Добавим к этому, совокупность призм построена на квадратной решетке (см. рис. 4.2). С учетом этих факторов рассмотрим, какое значение имеет смещение концов капилляров. Пространственное расположение капилляров имеет следующие особенности: 1) артериальные и венозные концы соседних сосудов (ребер призмы) последовательно смещены на полдлины вверх (или вниз) [337], 2) кровотоки в противоположных капиллярах имеют обратные направления [261]. Как мы уже показали выше, за счет замедления движения эритроцита по капилляру распределение рО2 по длине капилляра имеет приблизительно линейный характер (см. рис. 4.4). За счет этого феномена обеспечивается линейное распределение рО2 по длине каждого из 4 капилляров. При переходе от одного сечения призмы кдругому приращения величины рО2 в парах противоположных по диагонали сосудов одинаковы, но противоположны по знаку. Вследствие этого создается приблизительно равное распределение рО2 во всех сечениях систолических и диастолических призм. Потребление кислорода в течение систолы во всех сечениях «систолических» призм и в течение диастолы в «диастолических» одинаково. Таким образом, гармония «смещенного» сопряжения капилляров, каждый из которых занимает в призме оптимальное положение, обеспечивает одинаковое «качество» кислорода по всей высоте призмы (см. рис. 4.5, А). При любом другом взаимном смещении капилляров была бы нарушена аналогия оптимального распределения рО2 во всех сечениях призм.
4.5. Распределение рО2 в капиллярно-тканевыx призмах
В режимах гипертензии
Рассмотрим распределение рО2 в капиллярно-тканевых призмах в золотом и других режимах гипертензии. В заданном режиме гипертензии потребление кислорода единичным объемом мыщцы в каждый момент кардиоцикла увеличивается в b раз. Соответственно, плотность капилляров, открытых в систолу и диастолу, также возрастает b раз, что соответствует сокращению расстояния между сосудами в  раз и, соответственно, уменьшению объема призм в b раз. При заданном уровне гипертензии b систолическая призма А превращается в систолическую призму Б (см. рис. 4.5). Очевидно, что потребление кислорода «гипертензивной» призмой Б равно потреблению исходной призмы А; это равенство сохраняется при всех уровнях гипертензии. Исходя из (4.27), та же аналогия имеет место и для всех диастолических призм.
раз и, соответственно, уменьшению объема призм в b раз. При заданном уровне гипертензии b систолическая призма А превращается в систолическую призму Б (см. рис. 4.5). Очевидно, что потребление кислорода «гипертензивной» призмой Б равно потреблению исходной призмы А; это равенство сохраняется при всех уровнях гипертензии. Исходя из (4.27), та же аналогия имеет место и для всех диастолических призм.
|
| |  |
Рис. 4.5. Капиллярно-тканевая призма. А – в золотом, Б – в режиме гипертензии; l K (βЗС), l K ((β) - длина капилляра в золотом и других режимах гипертензии, l K ((β)=l K (βЗС); đ(βЗС), đ(β) - расстояние между капиллярами в золотом и других режимах гипертензии; đ(β)=đ(βЗС)/ Ö b. 
Потребление кислорода систолическими и диастолической призмами за единицу времени при любой гипертензии всегда одинаково и равно общей исходной величине в золотом режиме. Таким образом, сердечная мышца в любой момент времени при любой гипертензии состоит из «призм», каждая из которых за единицу времени потребляет одинаковое количество кислорода. Покажем особенности аналогии распределения рО2 по длине мышечного волокна в золотом режиме гипертензии. На протяжении отдельного мышечного волокна существует последовательность капиллярно-тканевых призм (см. рис. 4.6). Выберем в качестве исходного сечение одной из призм на уровне, соответствующем артериальному и венозному концам двух противоположных капилляров. При смещении от этого сечения на полдлины капилляра вниз распределение рО2 в данном сечении «поворачивается» на 90° относительно продольной оси волокна. При последующих смещениях вниз на 
полдлины капилляра исходное распределение «поворачивается» соответственно на 180о и 270о. При смещении вниз на 2 длины капилляра распределение рО2 совпадает с исходным. При последующем смещении через каждые 2lK цикл винтового «поворота» повторяется. Очевидно, что и при смещениях на углы в пределах 0o<y<90o, 90o<y<180o, 180o<y<270o, 270o<y<360o распределение рО2 от исходного отличается незначительно. Таким образом, распределение рО2 в поперечных сечениях мышечного волокна происходит в соответствии с винтовой симметрией. Такое пространственное сопряжение капилляров обеспечивает одинаковые условия для «качественного» кислородного обеспечения кардиомиоцитов по всей длине мышечного волокна. Благодаря этому обстоятельству в золотом режиме гипертензии ни один из кардиомиоцитов,составляющих мышечное волокно, не находится в менее благоприятных «кислородных» условиях, чем остальные. Гармония «противоположностей» в распределении концов капилляров по длине мышечного волокна создает условия
оптимального распределения величин рО2 по всей длине мышечного волокна. В конечном счете, форма капилляров, параллельность и винтовая симметрия их пространственного расположения по отношению к мышечным волокнам обеспечивают оптимальное сопряжение (гармонию) «качественного» и количественного обеспечения кислородом любого участка мышечного волокна и сердечной мышцы в целом.
Рассмотрим аналогию распределения рО2 в условиях гипертензии. Выберем для сравнения некоторые k-сечения призмы А (покой) и призмы Б (гипертензия), равноудаленные от основания (см. рис. 4.1 и 4.5). Распределение плотности рО2 в k-сечении призмы А соответствует некоторой кривой I (см. рис. 4.7). В капиллярах призмы Б k-сечении напряжение кислорода при гипертензии b возрастает на величину dpO2=42lgb. Поскольку потребление кислорода в обоих k-сечениях одинаково, то кривая распределения рО2 в k-сечении призмы Б смещается вправо на величину dpO2 (см. рис. 4.7, кривая II). Очевидно, что аналогичное смещение распределения рО2 происходит и в других сечениях призмы Б. С физиологической точки зрения сдвиг кривых распределения в сторону больших значений имеет важное значение. Установлено [289], что при снижении в кардиомиоцитах средней величины рО2 ниже 3,4-3,6 мм рт. ст. начинается фибрилляция сердца, свидетельствующая о кислородной недостаточности. Параллельный сдвиг вправо позволяет вывести клетки из летальной зоны (при росте гипертензии эта зона расширяется в сторону более высоких рО2).
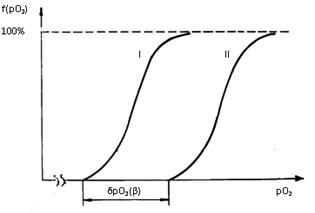
Рис. 4.7. Плотность распределения величин рО2 в k–сечении капиллярно-тканевой призмы [ 193 ]. По оси абсцисс – рО2 (условные единицы); по оси ординат – долевое распределение рО2 в k-сечении призмы (в %). I – распределение рО2 в золотом, II – распределение рО2 в некотором режиме гипертензии; δpO2(β)=42lgβ – приращение рО2 на концах капилляра в режиме гипертензии.
Поскольку при гипертензии b в каждом сечении призм и капилляров приращения dpO2(b) одинаковы, то постоянна и разность напряжений кислорода между кровью и тканью. Как следствие, постоянна и скорость отдачи кислорода движущимся эритроцитом [291]. Эта скорость неизменна по всей длине открытого сердечного капилляра в любой момент кардиоцикла в покое и при любой гипертензии. Она имеет, как было показано в 3 главе (см. раздел 3.6), для любого поперечного сечения каждого капилляра максимально возможное значение:
U( b) = 220%/c.
Эта величина является кислородным инвариантом всего капиллярного русла сердца. Механизм «винта» обеспечивает в каждом сечении мышечного волокна одинаковое «количество» и «качество» кислорода, адекватное заданному уровню гипертензии.
Подытожим. На примере тканевой призмы с ребрами-капиллярами нами проанализированы оба аспекта кислородного обеспечения клеток - по «количеству» и «качеству». Первый параметр в условиях гипертензии обеспечивается увеличением плотности капиллярной сети в b раз, второй - возрастанием линейной скорости эритроцитов в капиллярах также в b раз. Оба параметра связаны с увеличением среднего аортального давления в b раз. Можно сказать, Природа из диапазона возможных величин плотности капилляров, их взаимного расположения и возможных скоростей эритроцитов в капиллярах «выбрала» «рабочие» величины, оптимальные для каждого уровня гипертензии. Рабочие значения этих параметров разделяют диапазон допустимых значений на «противоположности». При всяком уровне гипертензии существует оптимальная гармония «противоположностей» для каждого из этих параметров. Это сопряжение позволяет обеспечить энергооптимальное снабжение кислородом любого региона сердечной мышцы как в покое, так и при всяком уровне гипертензии.
Следует отметить инварианты, присущие капиллярно-тканевым призмам в течение всего сердечного цикла в золотом и любом другом уровне гипертензии:
1) постоянство скорости отдачи кислорода из каждого открытого капилляра;
2) постоянство разности рО2 на артериальном и венозном концах капилляров;
3) постоянство «винтового» распределения рО2 вдоль мышечного волокна;
4) постоянство потребления кислорода объемами систолических и диастолических призм за единицу времени.
Выше нами были рассмотрены полностью «капиллярные» призмы (все ребра - капилляры). Рассмотрим призму, у которой артериола - одно из ребер. В сущности, как поставщики кислорода обменные артериолы почти ничем не отличаются от капилляров. Отметим ряд факторов архитектоники и гемодинамики, аналогичных для артериол и капилляров. Как мы уже отмечали раньше, стенки мельчайших артериол имеют строение, весьма сходное со строением капилляров: эпителий, тончайшая базальная мембрана и сравнительно редкие и очень тонкие пучки гладких мышц. Все эти образования имеют такой же коэффициент диффузии для О2 и СО2, как и окружающие ткани [66, т. 3, с. 182]. Обменные артериолы, также как и капилляры, параллельны мышечному волокну, которое они обеспечивают кислородом [270, 55]. Показано также [46], что при естественном отношении S/V энергия деформации мембраны эритроцитов минимальна. Вследствие этого свойства изменение формы эритроцитов в микрососудах происходит с минимумом потерь энергии. Эритроциты при перемещении в артериолах с диаметром менее 40 мкм так же, как и в капиллярах, принимают парашютообразную форму [326], которая обеспечивает оптимальные условия для отдачи кислорода [292]. В артериолах, как и в капиллярах, отношение средних скоростей движения эритроцитов и плазмы приблизительно равно 1,6, т. е. золотому числу [230, 282]. Как нами показано выше, каждое сечение артериол, независимо от их диаметра, отдает за единицу времени то же количество кислорода, что и сечение капилляра. Обьем отданного кислорода той или иной артериолой за единицу времени зависит лишь от ее длины, но не от диаметра. С «кислородной» точки зрения артериола фактически тот же капилляр. Таким образом, «включение» артериол в призму вместо капилляров ничего не меняет за исключением одного обстоятельства – величины рО2 на концах артериол значительно больше, чем на концах капилляров. Однако в тканях и это различие нивелируется по мере удаления от стенки сосудов. Показано [247], что чем больше диаметр артериол, тем менее они проницаемы для кислорода, тем больше падение рО2 на стенке. У капилляров при небольшом удалении от стенки сосуда снижение тканевого рО2 по отношению к крови незначительно. В то же время при таком же удалении от стенки артериол величина рО2 резко снижается, почти достигая «капиллярных» значений [71]. При дальнейшем удалении от стенки снижение рО2 происходит по аналогии с капиллярами. Следовательно, «вставка» артериолы вместо капилляра в плане адекватного обеспечения кислородом по «качеству» и «количеству» тканей практически ничего не меняет – распределение рО 2 в любом сечении призм во всех режимах гипертензии сохраняется таким же, что и при «капиллярном» варианте. Однако между артериолами и капиллярами есть существенное различие. Разница состоит в «цене» потребляемого из них кислорода: энергетическая «стоимость» единичного объема кислорода, потребляемого из артериол, выше, чем в капиллярах.
Заключение. Гармония оптимального сопряжения «противоположностей» архитектонических, гемодинамических и кислородных параметров в системе «Капиллярно-тканевая призма» обеспечивает оптимальное кислородное обеспечение последней по «количеству» и «качеству» с минимальными затратами энергии, крови и сосудистого материала во всех режимах гипертензии.