Система крово-кислородного обеспечения сердечной мышцы имеет важнейшие значение для обеспечения нормальной деятельности сердца и всего организма. Эта система имеет «цель»: оптимальное сопряжение «количества» и «качества» кислорода, потребляемого в сердечных клетках. Отметим, сердце человека и других млекопитающих в покое потребляет около 10% от общего количества кислорода, потребляемого организмом, хотя вес сердца составляет всего около 0,6% от веса тела. Энергию, необходимую для совершения механической работы, сердце получает главным образом за счет распада углеводов и жиров с участием кислорода. При интенсивной мышечной работе коронарный кровоток может возрастать в четыре раза; примерно во столько же раз увеличивается потребление сердцем кислорода [193].
Математиками представлены несколько моделей кислородного обеспечения сердечной мышцы. Однако все существующие к настоящему времени математические модели страдают существенными недостатками. Даже в наиболее продвинутых моделях [261] не представлено сопряжение кислородных параметров в сердечной ткани с параметрами архитектоники и гемодинамики снабжающих микрососудов. Отсутствие этих параметров не позволяет в полной мере представить их вклад в сложную систему крово-кислородного обеспечения. Нашей задачей является выявить с учетом не рассматриваемых прежде параметров конечную «цель» Природы: «Почему эта система организована таким, а не иным образом?». Для решения этой задачи ниже нами произведена последовательная «сборка» системы кислородного обеспечения сердца по направлению «от простого к сложному». «Сборка» докапиллярного участка произведена по цепочке функционально связанных кислороднесущих систем возрастающей сложности: эритроцит → кровь → сосуд с движущейся кровью → гемососудистый тройник → генерации транспортных и обменных тройников → докапиллярное русло сердца. На каждом этапе во всех рассматриваемых системах будет представлена гармония «противоположностей».
Эритроцит
Первой по «простоте» кислороднесущей системой является эритроцит. Для всех позвоночных универсальным признаком является заключение гемоглобина в специальные клетки – эритроциты. Молекулы гемоглобина обладают способностью присоединять и отдавать молекулярный кислород. Основная масса эритроцита состоит главным образом из воды (65%) и молекул гемоглобина (34%). Отметим, что соотношение масс гемоглобина и воды близко к соотношению чисел Фибоначчи (1:2). Концентрация гемоглобина у разных видов млекопитающих одинакова; она не зависит от диаметра и объема эритроцитов и их возраста [88, 171].
«Вхождению» гемоглобина в эритроцит соответствует ряд конструктивных особенностей, аналогичных для различных видов млекопитающих:
1) оболочка эритроцита отделяет дыхательный пигмент от плазмы крови, что создает наиболее благоприятную ионную среду для выполнения клеткой специфических кислороднесущих функций;
2) внутри эритроцита концентрация гемоглобина достигает достаточно высокой степени, что необходимо для сохранения пигментом сложного строения, чтобы обеспечить ему кооперативный эффект и своеобразную форму кривой диссоциации оксигенированной его формы;
3) высокая концентрация гемоглобина внутри эритроцита и изоляция пигмента от плазмы позволяет резко повысить кислородную емкость крови без увеличения онкотического давления в плазме, что имеет огромное значение для обмена воды и электролитов между тканями и кровью в тканевых капиллярах;
4) кооперативное связывание кислорода повышает эффективность гемоглобина как переносчика кислорода [152];
5) процессы связывания и отделения кислорода гемоглобином не требуют энергетических затрат [171].
6) молекулы гемоглобина в эритроците имеют паракристаллическую упаковку; их концентрация в клетке близка к предельной [218].
Представленные особенности «включения» гемоглобина в систему «эритроцит» обеспечивают последнему эффективное взаимодействие с окружающей средой. Прежде всего, нахождение молекул гемоглобина внутри эритроцита обеспечивает по сравнению с растворением такого же количества гемоглобина в плазме уменьшение вязкости крови в несколько раз [188]. Естественной концентрации гемоглобина в крови (15 г на 100 мл) соответствует минимальная вязкость последней; даже небольшое увеличение концентрации гемоглобина резко увеличивает вязкость крови [214]. Эритроциты придают крови свойства неньютоновской жидкости, что обеспечивает значительное снижение вязкости при прохождении крови через цепочку микрососудов.
Вода представляет собою двойную структуру. Одна ее фракция имеет квазикристаллическую структуру, вторая – это обычная жидкая вода. Физическая особенность внутриклеточной воды заключается в ее упорядоченной, квазикристаллической структуре при одновременном сохранении свойства жидкой воды – низкого значения вязкости. Внутри клетки квазикристаллическая вода находится в состоянии максимальной упорядоченности – в состоянии, достигаемом в неживых системах только при абсолютном нуле. Это состояние связано с тем, что квазикристаллическая структура внутриклеточной воды при температуре около 38оС непрерывно разрушается и непрерывно возникает заново в процессе расходования энергии метаболизма [171], т.е. происходят непрерывные фазовые переходы «противоположностей» (кристалл – жидкость) [171].
Добавим к вышесказанному, что вода - супрамолекулярная аквасистема, межмолекулярный континиум которой обеспечен единой динамической Н-системой водородных связей [146]. В аквасистемах клетки, ткани, органа имеет место оптимальное соотношение энергий организованных I(U) и хаотических S(U) форм движения. Вода способна сохранять уровень организованности своих фрагментов. На уровне клетки вблизи 38оС организованной формой является жидкий лед, жидкая вода соответствует хаотической форме. Во всех аквасистемах здорового организма существует поддерживаемый уровень организованности i=I(U)/S(U), равный золотому числу i=1,62 [54]. Таким образом, энергии «противоположностей» (организованность и хаос) в аквасистемах здорового сердца соотносятся между собою по золотому сечению! Отметим, максимальная способность воды переходить в «противоположные» состояния в обоих направлениях соответствует «золотой» температуре 38оС. Эта величина делит температурный диапазон воды по пропорции золотого сечения; при этой температуре создаются наиболее благоприятные условия для протекания процессов жизнедеятельности. Все неводные компоненты живой клетки (белки, нуклеиновые кислоты и т.д.) при обеспечении «квазикристаллической» температуры воды обладают термостабильностью при обмене веществ. Высокие теплофизические свойства воды обеспечивают стабильность среды в клетках и постоянство протекания биохимических процессов в последних при нормальных температурах в организме теплокровных животных. Таким образом, золотой уровень организованности «совмещен» с золотым разделением температурного интервала воды.
Молекулы гемоглобина располагаются на внутренней стороне мембраны эритроцита; между двумя молекулами гемоглобина находятся две молекулы воды. Вода в этой конструкции выполняет две взаимно «противоположные функции». С одной стороны, вода поддерживает жесткую структуру, что позволяет сохранять автономность каждой молекулы гемоглобина, с другой – обеспечивает изменчивость формы эритроцита. Равномерное распределение молекул гемоглобина в эритроците обусловливает равномерное распределение внутриклеточной воды. С другой стороны, внутриклеточная вода в эритроците образует сложную пространственную сеть и обладает, следовательно, квазикристаллической структурой. Энергия метаболизма безядерного эритроцита полностью расходуется для поддержания структуры внутриклеточной воды эритроцита в упорядоченном, квазикристаллическом состоянии [171].
Главная функция эритроцитов - транспорт кислорода к месту его потребления с последующим газообменом между кровью и окружающими тканями. Складирование гемоглобина под мембраной эритроцитов и эффективность движения последних в потоке крови обусловлены, прежде всего, особенностями формы эритроцитов. Оптимальная форма должна: 1) обеспечить при заданном объеме эритроцита «складирование» под мембраной максимального количества молекул гемоглобина; 2) обеспечить минимум потерь энергии при движении эритроцита в плазме крови. Этими двумя предварительными условиями обусловливается минимальная энергетическая «цена» транспорта кислорода.
Главная функция эритроцитов - транспорт кислорода к месту его потребления с последующим газообменом между кровью и окружающими тканями. Складирование гемоглобина под мембраной эритроцитов и эффективность движения последних в потоке крови обусловлены, прежде всего, особенностями формы эритроцитов. Оптимальная форма должна: 1) обеспечить при заданном объеме эритроцита «складирование» под мембраной максимального количества молекул гемоглобина; 2) обеспечить минимум потерь энергии при движении эритроцита в плазме крови. Этими двумя предварительными условиями обусловливается минимальная энергетическая «цена» транспорта кислорода.
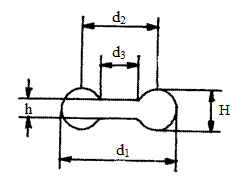
У всех млекопитающих от крошечной землеройки до кита эритроциты имеют форму двояковогнутого диска. Диаметр эритроцитов у всех млекопитающих приблизительно одинаков и составляет 7-10 мкм. Для нас представляет интерес отношение поверхности мембраны эритроцита, под которой размещаются молекулы гемоглобина, к объему клетки. При анализе возможных дискообразных форм эритроцита при постоянном объеме было показано, что максимальное отношение его поверхности S к объему V соответствует естественной форме (для человека S/V=1,638) [284]. Молекулы гемоглобина располагаются непосредственно под мембраной эритроцита; их взаимное расположение соответствует предельно плотной, паракристаллической упаковке [218]. Таким образом, эритроцит имеет: 1) максимальное отношение S/V, 2) максимально плотную упаковку гемоглобина под мембраной. Максимальная концентрация гемоглобина, равная 34%, наблюдается у эритроцитов разных видов млекопитающих, причем эта концентрация не зависит от диаметра эритроцитов и их возраста [171]. Эритроцит транспортирует максимальное количество молекул гемоглобина, а, следовательно, и молекул кислорода. Следовательно, «складские» возможности эритроцита использованы в высшей мере эффективно. За счет этого доставка необходимого количества кислорода к месту потребления достигается при минимальном количестве «суденышек» (эритроцитов). Установлено [166], что оптимальная конструкция эритроцитов связана с числами Фибоначчи: 1, 2, 3, 5, 8 (см. рис. 3.1)
| Рис 3.1. Дискообразная форма эритроцита человека. d1 =8 мкм- диаметр клетки; d2 =5 мкм- диаметр наиболее выпуклой части клетки; d3 =3 мкм- диаметр вогнутой части клетки; Н=2 мкм– толщина выпуклой части клетки, h=1 мкм - толщина клетки.
| |
.
Главная функция эритроцитов - транспорт кислорода к месту его потребления с последующим газообменом между кровью и окружающими тканями. Складирование гемоглобина под мембраной эритроцитов и эффективность движения последних в потоке крови обусловлены, прежде всего, особенностями формы эритроцитов. Оптимальная форма должна: 1) обеспечить при заданном объеме эритроцита «складирование» под мембраной максимального количества молекул гемоглобина; 2) обеспечить минимум потерь энергии при движении эритроцита в плазме крови. Этими двумя предварительными условиями обусловливается минимальная энергетическая «цена» транспорта кислорода.
Большое значение для гемодинамики имеют механические свойства мембраны эритроцита. Показано [46], что при естественной форме эритроцита энергия, необходимая для деформации мембраны минимальна. Благодаря свойствам растяжимости мембраны движение эритроцитов в плазме происходит с минимальными затратами энергии; эритроциты движутся подобно капелькам жидкости [254].
При движении эритроцита в потоке происходит вращательно-поступательное движение мембраны вокруг содержимого клетки (оболочка эритроцита движется подобно гусенице танка) [315]. Таким образом, оптимальная форма мембраны обусловливает максимальную эластичность последней и минимум потерь энергии при движении отдельного эритроцита. Отметим, что такие замечательные особенности мембраны присущи лишь эритроцитам, «построенным» по числам Фибоначчи!
Хаотическое расположение эритроцитов в крови исключается [95, 208]; каждый эритроцит движется вдоль сосуда своей узкой стороной. Отметим, что дискообразная форма эритроцитов препятствует их оседанию на стенки сосуда [76]. При таком расположении, находясь в слоях плазмы с разными скоростями, эритроцит вращается вокруг своего центра симметрии по оси, перпендикулярной к его вогнутым сторонам.
Система морфологических и корпускулярных элементов крови окутана силовыми линиями электрического поля. Между отдельными элементами крови действуют силы электростатического распора, препятствующие полному сближению и соприкосновению эритроцитов. Дискоидная форма эритроцитов обеспечивает симметрию их движения по отношению к продольной оси сосуда; благодаря этому обстоятельству обеспечивается минимальная вязкость крови [335]. Таким образом, дискоидная форма эритроцита, «построенного» в соотношениях Фибоначчи, является оптимальным разрешением высоких требований к гемодинамическим параметрам этой клетки и крови в целом. Можно сказать, что форма клетки найдена как оптимальное сопряжение «противоположностей» параметров в сложной системе (эритроцит). Отметим, что в капиллярах и терминальных артериолах эритроциты - диски принимают парашютообразную форму [292]. Эритроциты выстраиваются в цепочку, которая перемещается в оболочке из плазмы. Бесклеточная краевая зона с низкой вязкостью образует слой, который значительно облегчает скольжение эритроцитов. Этот феномен противодействует увеличению вязкости крови при уменьшении скорости кровотока. Установлено [285a], что вязкость крови в каждом сосуде не возрастает, если скорость эритроцитов не опускается ниже 500 мкм/сек. однако даже в капиллярах скорость значительно выше критической.
Отметим «золотые» особенности в конструкции молекулы воды. Эта молекула имеет форму в виде буквы V. Для состояния льда отношение длин связей (Н-О) к (Н-Н) равно 0,100/0,163=0,613, в талой воде (Н-О)/(Н-Н) приближается к значению j=0,618, т. е. имеют место «золотые» соотношения [13, 194]. «Золотое» расположение атомов обусловливает особые свойства воды среди других жидкостей. Такая конструкция сильно отличает молекулу Н2О от линейных молекул, в которой все атомы расположены цепочкой. Вода – единственная жидкость на Земле (кроме ртути), для которой зависимость удельной теплоемкости от температуры имеет минимум (см. рис. 3.2). Именно такое особое расположение атомов в молекуле воды и позволяет ей иметь множество необычных свойств. Вода внутри клетки находится в состоянии максимальной упорядоченности, достигаемой в неживых системах только при абсолютном нуле. Отметим, что теплоемкость воды при золотой температуре 38оС незначительно отличается от минимальной при температуре 36,8оС. Вода при температуре 38оС в клетках обладает особой, квазикристаллической упорядоченностью, при которой количество разорванных и сохранившихся тетрамерных связей примерно одинаково [87]. При «золотой» температуре 38оС выравниваются массы квазикристаллической
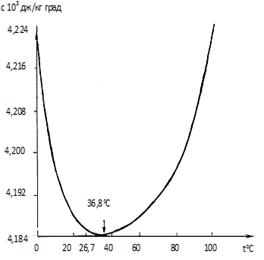
Рис. 3.2.Зависимость теплоемкости воды от температуры
[ 23 ]. Интервал toC = 26,7 – 43 o C соответствует
интервалу жизни биосистем.
воды (жидкий лед) и обычной воды, при этом способность воды переходить из одного состояния в другое (т.е. обмен информацией) максимальна. Такое соотношение обеспечивает равновероятное протекание биопроцессов в обоих направлениях. Температура 38оС обеспечивает в эритроците условия теплового комфорта, в условиях которого на поддержание оптимальной температуры в клетке расходуется минимальная энергия (см. рис. 3.2). Вблизи этой температуры кровь при гематокрите Ht=42% имеет минимальную вязкость [166]. Температура на участке 36,8 - 38,2оС оптимальна для течения ферментативных реакций [53]. При превышении температуры 38,2оС скорость ферментативных реакций начинает снижаться вследствие разрушения вторичных и третичных структур фермента. Таким образом, «золотая» температура является рабочей температурой, которая удовлетворяет, как мы могли убедиться, оптимальной гармонии «противоположностей» многих параметров не только эритроцита, но и крови в целом.
В большинстве «простых» систем эритроцита имеют место золотые «противоположности». Оптимальное «вхождение» «противоположностей» каждого из гемодинамических и структурных, а также температурного, параметров в систему «Эритроцит» обеспечивает свой «вклад» в максимальную экономию энергии и вещества в системе «Кровь».
Кровь
Основными элементами крови являются плазма, эритроциты, лейкоциты и тромбоциты. Два последних элемента не имеют отношения к транспорту кислорода, поэтому мы не будем их рассматривать. Будет произведен анализ «вхождения» в кровь первых двух элементов: эритроцитов и плазмы.
Оптимальная величина концентрации эритроцитов в крови обусловлена двумя противоположными требованиями. С одной стороны, желательно иметь высокую концентрацию эритроцитов, поскольку при этом возрастает количество кислорода в единичном объеме крови. С другой стороны, неограниченный рост гематокрита связан с повышением вязкости крови, а, следовательно, и с возрастанием расхода механической энергии на транспорт кислорода. Это противоречие устраняется [203, 293], если отношение вязкости крови ξ к гематокриту Ht минимально (см. рис. 3.3). Такое оптимальное отношение соответствует естественной величине Ht=42% при нормальном артериальном давлении и нормальном напряжении сдвига τ=103 дн/см2 [242]. По расчетам И.Ф. Образцова и М.А. Ханина [113] величине Ht=42% соответствует оптимальная концентрация эритроцитов С=5,2·109 1/мл крови. Экспериментально установлено [246], что минимальная «цена» транспорта единичного объема кислорода в сосудах приблизительно соответствует естественной величине гематокрита. Гематокрит, определенный путем центрифугирования порции крови из артериол и венул диаметром 100-200 мкм, равен постоянной величине, которая почти не отличается от гематокрита в магистральных сосудах [229, 281]. Таким образом, оптимальное соотношение «твердой» (эритроциты) и «жидкой» (плазма) составляющих крови остается постоянным по всему коронарному руслу. «Оптимальная» величина гематокрита одновременно обусловливает оптимальность реологических свойств крови и объема транспортируемого кислорода. «Рабочая» величина этого параметра оптимальным образом разделяет возможный диапазон вязкости крови и возможный диапазон концентрации эритроцитов. Вследствие оптимальной гармонии «противоположностей» этих параметров на транспорт единичного объема кислорода по коронарному руслу тратится минимальное количество энергии. Установлено [281], что естественное соотношение объемов эритроцитов и плазмы соответствует отношению средней скорости эритроцитов к средней скорости пристеночного слоя плазмы, равном 1,6 (золотое число!). «Золотое» соотношение скоростей эритроцитов и плазмы, соответствующее оптимальному соотношению гематокрита и вязкости крови, обеспечивает согласованное по времени движение «твердых» и «жидких» компонентов крови в каждом сосуде. Кровь млекопитающих на 80% состоит из воды. Отметим, потребность в энергии, расходуемой на поддержание температуры самого теплоносителя при 38°С минимальна [166]. При этой температуре выравниваются массы квазикристаллической воды (жидкий лед) и обычной воды [87]. Этот феномен обеспечивает равновероятность течения обратимых биохимических реакций в обе стороны. Добавим к этому, вода имеет высокую теплопроводность и теплоемкость. Благодаря первому свойству тепло от химических реакций равномерно распределяется по объему крови (при этом устраняется «перегрев» и «недогрев»); вследствие большой теплоемкости даже для небольшого повышения температуры требуется большое количество энергии. Тем самым тепловые свойства воды обеспечивают постоянство протекания биохимических процессов в крови и ее температурную стабильность. Как отмечали Эйген и Винклер (220, с. 96), «в живых организмах скорость химических реакций всегда достигает предельного значения, которое определяется законами физики». При «золотой» температуре 38оС в плазме крови существуют наиболее благоприятные условия для протекания биологических процессов с максимально возможной скоростью.
Подытожим. Минимизация затрат механической энергии на перемещение крови происходит благодаря минимизации вязкости последней (оптимальная форма и оптимальная концентрация эритроцитов, симметрия кровотока, оптимальная температура). С другой стороны, за счет оптимальной формы эритроцитов и паракристаллической упаковки молекул гемоглобина обеспечивается максимально возможная концентрация дыхательного пигмента в единичном объеме крови. Оптимальное «вхождение» «противоположностей» каждого из этих параметров в систему «Кровь» обеспечивает минимум потерь механической энергии на перемещение единичного объема кислорода к месту его потребления. Естественно, что все представленные оптимальные «рабочие» значения параметров одновременно характеризуют реологические свойства крови и объем транспортируемого кислорода. Естественно, все рассмотренные нами особенности вхождения «противоположностей» в систему «Кровь» имеют место во всех органах организма (в том числе и в сердце).
Интересно добавить, что установлена связь элементов крови с золотой пропорцией и числами Фибоначчи [145]. Форменные элементы крови - «кислородные» (эритроциты) и «некислородные» (лейкоциты и тромбоциты) - в глобулярном объеме крови сбалансированы как соотношение чисел 62:32:6. Нетрудно увидеть, что отношение «противоположностей» - «кислороднесущей» и «некислородной» - равно золотому числу 1,618. Можно сказать, что и здесь налицо золотая гармония «противоположностей» в элементах, несущих кислород, и элементах, имеющих «некислородное» назначение. Было также показано отношение «противоположностей»: объема циркулирующей крови к плазматическому объему в организме равно 5:3, а отношение плазматического объема к глобулярному – 3:2 (числа Фибоначчи). Кроме того, А.Г. Суббота [166] показал, что соотношение «противоположностей» - эритроцитов трансформированной и дискоидной форм - также приблизительно составляет золотое сечение - 38,2 и 61,8%. С большой степенью вероятности можно предположить, что и за этими «золотыми» соотношениями скрываются дополнительные аспекты экономии энергии и вещества.
Сосуд с движущейся кровью
В сосудах с движущейся кровью расход энергии можно разделить на «механическую» и «химическую» составляющие. Механическая энергия тратится на перемещение крови в сосуде, а «химическая» расходуется на биопроцессы в крови. Рассмотрим механическую составляющую. Выше нами были представлены общие пути оптимизации вязкости крови: «подбор» формы эритроцитов, гематокрита и температуры. Однако в сосуде имеют место дополнительные факторы минимизации вязкости, связанные с движением эритроцитов. Дискообразная форма эритроцитов обеспечивает симметричное движение последних относительно продольной оси сосуда, что создает дополнительное снижение вязкости крови [335]. Таким образом, минимальная величина вязкости крови в сосуде обусловлена не только оптимальным «подбором» гематокрита, температуры и формы эритроцитов, но также и симметрийным способом движения последних. Наблюдения показали высокоупорядоченное распределение клеточных и плазменных компонентов в крови. Более легкие и мелкие компоненты крови находятся у стенок сосудов, а самые тяжелые смещены к продольной оси; тем самым обеспечиваются минимальные потери энергии на трение потока у стенок сосудов. Существуют и другие аспекты экономии механической энергии в сосуде. Установлено [94, 95], что движение крови в артериях имеет ламинарно-спиральный характер (можно провести некоторую аналогию с движением пули в стволе с винтовой нарезкой). Движение такого рода обеспечивается спиралевидной ориентацией мышечных волокон в стенках артерий [94]. Спиральный тип движения универсален и происходит c минимальной потерей энергии [43, 290]. Давно известно, что спирализация конструкций и процессов способствует экономии энергии и материала, обеспечивает повышенную прочность системы и надежность ее функционирования. Установлено также соответствие ориентации эндотелиальных клеток протекающему кровотоку [288]. Таким образом, налицо различные аспекты минимизации «механической» части общей энергии, расходуемой в сосуде (гематокрит, температура, симметрирование кровотока и спирально-ламинарный характер движения). Отметим также, что в сердечных сосудах имеют место синхронные пульсации стенки сосудов и кровотока. Частоты пульсации в стенке сосуда и потоке крови совпадают, между ними возникает резонанс. Резонанс между стенкой и кровью обеспечивает дополнительную минимизацию затрат энергии по перемещению крови в сосудах [173]. Таким образом, при движении крови в сосудах за счет рассмотренных аспектов обеспечивается дополнительная экономия механической энергии.
Необходимо рассмотреть соотношение затрат механической и «химической» мощностей при постоянном кровотоке в сосуде при изменении диаметра последнего.
Механическая мощность, расходуемая в сосуде с движущейся кровью (см. рис. 3.4),
W M = 128 Q 2 ξl / πd 4,
где Q – кровоток, ξ – вязкость крови, l, d – длина и диаметр сосуда.
«Химическая» мощность, потребляемая в сосуде:
W X = ( π /4) bd 2 l,
где b– расход химической энергии в единичном объеме крови за единицу времени.
Рис. 3.4. Сосуд с движущейся кровью. q - объемный кровоток, d , l - диаметр и длина сосуда, ξ - вязкость крови, b - удельный расход химической энергии за единицу времени.
Даже при первом взгляде на эти формулы очевидно, что механическая и химическая составляющие суммарной мощности зависят от диаметра в различной степени. Более того, при изменении величины диаметра они изменяются в противоположных направлениях. Следовательно, при заданной величине кровотока q минимальный суммарный расход «противоположных» мощностей - механической и химической - должен соответствовать некоторому оптимальному диаметру d. Режим движения кровотока в сосуде при минимальном суммарном расходе мощности получил обозначение - режим «минимальной работы» [295].
В золотом режиме гипертензии с учетом диаметра d(bЗС), длины сосуда l(bЗС) и времени нахождения эритроцитов в сосуде t(bЗС) объемный кровоток q(bЗС) можно представить следующим выражением:
q ( b ЗС ) = ( π /4) d ( b ЗС )2 l ( b ЗС )/ t ( b ЗС ).
В соответствии с расчетами Мюррея в этом режиме объемный кровоток в сосуде связан с его оптимальным диаметром и параметрами крови [295]:
q ( b ЗС )/ d ( b ЗС )3 = ( π /32) 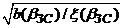 ,
,
где b(bЗС) – расход химической энергии в единичном объеме крови за единицу времени, x(bЗС) - вязкость крови в золотом режиме гипертензии.
Минимизация суммарного расхода энергии имеет место, если
/8 = l ( b ЗС )/ d ( b ЗС ) t ( b ЗС ).
Отметим, энергооптимальное сопряжение «противоположных» мощностей имеет место во всех сердечных сосудах [216, 203].
Оптимальное соотношение механической и химической мощностей равно постоянной величине для всех сердечных артерий, артериол и капилляров не только в покое, но и при различных уровнях физической нагрузки [203]:
g( bЗС) = W M( bЗС)/ W X( bЗС) = W M( b)/ W X( b)= 0,5.
Таким образом, энергооптимальная гармония «противоположных» мощностей во всех сердечных сосудах представлена постоянным числом. Оптимальное сопряжение «противоположных» мощностей обусловлено энергооптимальными «противоположностями» в архитектонических, гемодинамических, температурных и «химических» параметрах. Вследствие этого, энергетическая «цена» транспортируемого кислорода в каждом сердечном сосуде имеет минимальную величину. Добавим к этому, что механическая и химическая «противоположности» соотносятся между собою как числа Фибоначчи (1:2).
3.3. Бифуркация – оптимальный вариант ветвления докапиллярных сосудов
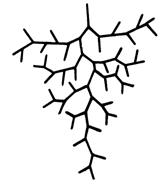
Бифуркация является общим типом разветвления для всех коронарных артерий, включая и артериолы [358]. Любая бифуркация (тройник) является «сложной» системой, в которую ствол и каждая из ветвей включены в качестве «простых» систем. Особенность конструкции тройника состоит в том, что каждая его ветвь является стволом следующего тройника. Совокупность тройников образует артериальное «дерево» (см. рис. 3.5).
Докапиллярное русло сердца (артериальное «дерево») включает в себя сосуды эластического и мышечного типа, а также микрососуды. Диаметр эластических артерий превышает 500 мкм. Сосудами мышечного типа принято считать артерии с диаметром от 100 до 500 мкм. В обменные генерации входят микрососуды - артериолы с диаметром менее 100 мкм [111]. По отношению к транспорту и потреблению кислорода докапиллярное русло можно разделить на два «противоположных» участка. Первый участок составляют эластические и мышечные сосуды; по этим сосудам осуществляется транспорт кислорода к обменному участку. На втором, микрососудистом, участке происходит диффузия кислорода в окружающие ткани. Транспортные сосуды сердца (d>100 мкм) всегда разветвляются на асимметричные ветви. Обменные микрососуды, напротив, имеют симметричное ветвление. Особенности архитектоники бифуркационных тройников в золотом режиме гипертензии можно представить следующими коэффициентами [216]:
1) коэффициент ветвления
k( b ЗС) = [ d 1( b ЗС)2 + d 2( b ЗС)2]/ d С( b ЗС)2,
2) коэффициенты относительного сечения ветвей
H 1( b ЗС) = d 1( b ЗС)2/[ d 1( b ЗС)2 + d 2( b ЗС)2],
H 2( b ЗС) = d 2( b ЗС)2/[ d 1( b ЗС)2 + d 2( b ЗС)2],
Из этих сотношений следует [193]:
d 1( b ЗС) =  d С( b ЗС), (3.1)
d С( b ЗС), (3.1)
d 2( b ЗС) = 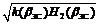 d С( b ЗС). (3.2)
d С( b ЗС). (3.2)
Отметим, что для сердечных тройников величина k(bЗС) находится в пределах 1<k(bЗС)£1,26 (режим «минимальной работы») [216]. В диапазоне 1<k(bЗС)£1,26 имеет место оптимальное сопряжение диаметров ствола и ветвей [131]:
d 1( b ЗС)3+ d 2( b ЗС)3= d С( b ЗС)3. (3.3)
Соотношение (3.3) соответствует минимальному расходу мощности, израсходованной в каждом сосуде тройника на диссипацию кровотока и биохимические процессы в крови [131].
Представим доказательства того, что бифуркация является оптимальным вариантом ветвления сердечных сосудов по сравнению с трифуркацией, тетрафукарцией и т. д. Рассмотрим в качестве сравнения наиболее близкие варианты ветвлений - бифуркацию (ствол и 2 ветви) и трифуркацию (ствол и 3 ветви). Предварительно примем равные условия для обоих вариантов:
1) одинаковая разность давлений между аортой и артериальным концом капилляров;
2) равенство кровотока в первой коронарной артерии;
3) одинаковый диаметр первой коронарной артерии и одинаковый диаметр капилляров;
4) одинаковая вязкость крови во всех сосудах;
5) одинаковая плотность капиллярной сети;
6) в стволе и ветвях бифуркационных и трифуркационных тройников имеет место оптимальный режим «минимальной работы»;
7) суммарные величины расхода энергии, объема крови в сосудах и проводимости на докапиллярном участке одинаковы для обоих вариантов.
Для упрощения расчетов примем на докапиллярном участке все бифуркационные и трифуркационные разветвления симметричными и вязкость крови во всех сосудах одинаковой. Коэффициент конструкции всякого сосуда для обоих видов разветвления будем также считать одинаковым (C=l/d =const).
Исходя из условия C=l/d =const и условия энергооптимальности для каждого трифуркационного разветвления в золотом режиме гипертензии имеем:
d 1( b ЗС)3+ d 2( b ЗС)3+ d 3( b ЗС)3= d С( b ЗС)3, (3.4)
l 1( b ЗС)3+ l 2( b ЗС)3+ l 3( b ЗС)3= l С( b ЗС)3, (3.5)
где dС(bЗС), d1(bЗС), d2(bЗС), d3(bЗС), lC(bЗС), l1(bЗС), l2(bЗС), l3(bЗС) - соответственно диаметр и длина ствола и 1, 2 и 3 ветвей.
При симметричном бифуркационном ветвлении
d 1( b ЗС) = d 2( b ЗС) = 0,794d С( b ЗС), (3.6)
l 1( b ЗС) = l 2( b ЗС) = 0,794 l С( b ЗС). (3.7)
Соответственно, при симметричной трифуркации
d 1( b ЗС) = d 2( b ЗС) = d 3( b ЗС) = 0,694d С( b ЗС), (3.8)
l 1( b ЗС) = l 2( b ЗС) = l 3( b ЗС) = 0,694 l С( b ЗС). (3.9)
Покажем преимущества бифуркации на примере сердца собак. При бифуркации общее число докапиллярных генераций сердца N=28 [194]. Исходя из условий 1) – 3) и зависимости (3.8) при трифуркации для собак общее число докапиллярных генераций должно было бы равняться 18; по сравнению с бифуркацией количество этих генераций уменьшается в 1,6 раз. В соответствии с условиями (1) - (7), общий кровоток, общий объем крови и общая проводимость бифуркационных и трифуркационных сосудов, начиная от аорты до капилляров, принимаем равными. Для выполнения этих условий давление на каждом «трифуркационном» сосуде и его длина должны возрасти в 1,6 раза. При таком условии суммарный объем крови и суммарная проводимость для обоих вариантов ветвления будут равными. Однако при этом плотность обменных микрососудов в бифуркационном варианте больше, чем в трифуркационном, что имеет большое значение для наиболее эффективного кислородного обеспечения миокарда. Рассмотрим на примере сердца собаки, как осуществляется это преимущество. При соблюдении условия 5) количество обменных сосудов в каждой «одноименной» m-генерации (начиная от капилляров) при бифуркации будет больше, чем при трифуркации. К этому необходимо добавить, количество «трифуркационных» генераций в пределах кислородотдающего участка (7<d<40 мкм) всего 6 (при бифуркации - 9) [203]. Таким образом, в пределах диапазона 7<d<40 мкм при бифуркации имеет место наибольшее количество обменных микрососудов. Эти микрососуды совместно с капиллярами образуют общую обменную сеть максимальной плотности.
Рассмотрим друг







