ТРЕМАТОДОЗЫ ЖИВОТНЫХ
Ставрополь- Махачкала, 2011
УДК 619:616.99
Авторы: д.в.н., профессор Колесников В.И., д.в.н., профессор Атаев А.М., к.б.н. Газимагомедов М.Г.
Рецензент: д.б.н., профессор Алиев Ш.К.
д.в.н.. профессор Магомедов О.А.
ТРЕМАТОДОЗЫ ЖИВОТНЫХ
В данной монографии отражены вопросы общей характеристики класса трематод и их систематика. Описаны наиболее чаще встречаемые трематодозы с разбивкой их по видам животных, птиц и рыб. Для каждого заболевания подробно описаны возбудители с их рисунками, циклы развития, клиника и патологоанатомические изменения, диагностика, лечение и профилактика с учетом новых химиотерапевтических средств. Монография предназначена для студентов ветеринарных факультетов средних и высших учебных заведений, институтов переподготовки и повышения квалификации.
Общая характеристика трематод
Впервые о трематодах упоминается в середине XVII в. в работах известного итальянского ученого Ф. Реди, который описал сосальщика из печени коровы. В дальнейшем К. Линней объединил уже известных к тому времени 40 видов трематод в один род Fasciola. Первая капитальная сводка о трематодах принадлежит перу К. Рудольфи (1819), который признан отцом современной гельминтологии. Он обосновал для трематод самостоятельный класс (Trematoda) и дал описание 220 видам сосальщиков, объединив их в 5 самостоятельных родов. Дальнейшее значительное развитие исследований по трематодам относится ко второй половине XIX в., когда были выполнены основополагающие работы Р. Лейкартом, К. Бэром, Я. Стеенструпом, А. Томасом, А. Лоосом, М. Брауном и др., посвященные фауне и жизненным циклам сосальщиков. Развитие следующего весьма продуктивного этапа в изучении различных аспектов трематод связано с именем академика К. И. Скрябина, который является создателем советской школы гельминтологии. Он вместе со своими учениками (В. Е. Судариковым, Е. Я. Башкировым, Д. Н. Антипиным, М. М. Белопольской, В. П. Коваль и др.) выпустил многотомное (свыше 20 томов) издание «Трематоды животных и человека». В этой сводке дается подробное описание свыше 3 тыс. видов и представлена современная система трематод. До настоящего времени данные работы лежат в основе современной систематики трематод и являются фундаментальной сводкой по мировой фауне этих паразитов. Значительный вклад в развитие отечественной трематодологии внесли В. А. Догель и его ученики (Б.Е. Быховский, И. Е. Быховская, Т. А. Гинецинская, и др.). Их перу принадлежат блестящие работы, посвященные жизненным циклам, биологии и экологии трематод. Так, Т. А. Гинецинской и ее учениками выпущены основополагающие работы, посвященные исследованиям жизненных циклов трематод. Интерес к трематодам не ослабевает и в наше время. С одной стороны, среди трематод имеется большое количество возбудителей опасных заболеваний, как человека, так и домашних животных. Данное обстоятельство является причиной развития прикладной трематодологии, связанной с разработкой в первую очередь медико-ветеринарных проблем. С другой стороны, трематоды с их уникальным жизненным циклом - это объект исследований, имеющих общезоологическую и общебиологическую направленность. В настоящее время российскими (В. Е. Судариков, А. А. Шигин, В. А. Ройтман, С. А. Беэр, Л. В. Филимонова, Т. А. Краснолобова, И. А. Добровольский, К. В. Галактионов и др.) и зарубежными (J. Pearson, K. Odening, K. Combes, D. Brooks, W. Haas, R.Wilson, Z. Zdarska и др.) учеными активно развиваются эти направления трематодологии. Класс Trematoda Rudolphi, 1808 - это большая группа гельминтов в составе типа плоских червей (Platyhelminthes). В настоящее время описано около 4000 видов сосальщиков. Трематоды ведут исключительно паразитический образ жизни, являясь эндопаразитами. Взрослые (половозрелые) трематоды - это, как правило, гермафродитные организмы, которые называются маритой. Раздельнополыми являются лишь представители семейства Schistosomatidae. Трематоды, главным образом, паразитируют у позвоночных животных, гораздо реже встречаются у беспозвоночных. Мариты преимущественно обитают в различных органах пищеварительной системы хозяина (желудок, кишечник, печень, поджелудочная железа). Определенное число видов паразитируют и в других отделах и органах хозяина (легких, носоглотке, почках, мочевом пузыре, грудной и брюшной полостях, кровеносной системе и т.д.). Трематоды сравнительно часто регистрируются у животных и человека, вызывая заболения, именуемые трематодозами. Особенно широко трематодозы распространены в странах с теплым климатом. Одна из наиболее характерных особенностей трематод, в отличие от остальных классов плоских червей - сложный жизненный цикл.
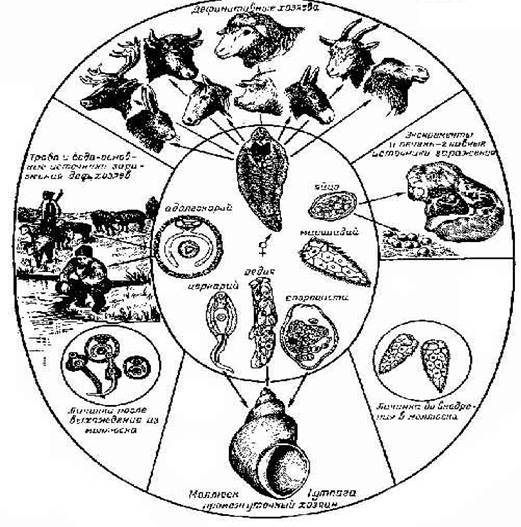
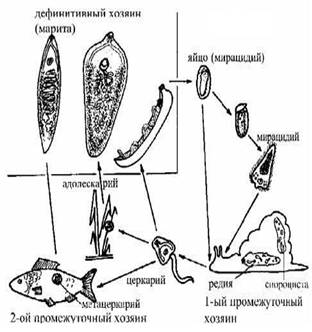 Основные этапы жизненного цикла. Хозяин, в котором обитают взрослые особи, именуется дефинитивным (окончательным). Хозяева, в которых размножаются и развиваются другие стадии цикла, называются промежуточными. Их может быть от одного до трех. Причем, первым (нередко единственным) промежуточным хозяином являются брюхоногие моллюски, вторым (если он есть) - беспозвоночные (кольчатые черви, моллюски, насекомые, ракообразные и др.) и позвоночные (рыбы, амфибии) животные. Жизненный цикл современных трематод - это чередование нескольких (минимум двух) самостоятельных поколений, различающихся, во-первых, по способу размножения (гермафродитное и партеногенетическое) и, во-вторых, по приуроченности к тем или иным животным-хозяевам. Взрослые трематоды, паразитируя в организме хозяина, продуцируют яйца, которые попадают во внешнюю среду. В яйце развивается мирацидий (личинка материнской спороцисты) - первая личиночная стадия в жизненном цикле трематод. Мирацидий активно (самостоятельно внедряется) или пассивно (не выходит из яйца) попадает в моллюска. После заражения первого промежуточного хозяина (моллюска) в нем развиваются партеногенетические поколения - спороцисты и редии. Партениты в большинстве случаев представлены не одной, а несколькими генерациями, сменяющими друг друга. Завершающей стадией онтогенеза партеногенетических поколений является образование гермафродитного поколения трематод. В зародышевой полости дочерней партениты протекает морфогенез первой гермафродитной личинки - церкарии. Зрелые церкарии (похожи на микроскопических головастиков) выходят из моллюска и ведут активный образ жизни во внешней среде, как правило, водной. Затем они, либо инцистируются на субстрате и определяются как адолескарии
Основные этапы жизненного цикла. Хозяин, в котором обитают взрослые особи, именуется дефинитивным (окончательным). Хозяева, в которых размножаются и развиваются другие стадии цикла, называются промежуточными. Их может быть от одного до трех. Причем, первым (нередко единственным) промежуточным хозяином являются брюхоногие моллюски, вторым (если он есть) - беспозвоночные (кольчатые черви, моллюски, насекомые, ракообразные и др.) и позвоночные (рыбы, амфибии) животные. Жизненный цикл современных трематод - это чередование нескольких (минимум двух) самостоятельных поколений, различающихся, во-первых, по способу размножения (гермафродитное и партеногенетическое) и, во-вторых, по приуроченности к тем или иным животным-хозяевам. Взрослые трематоды, паразитируя в организме хозяина, продуцируют яйца, которые попадают во внешнюю среду. В яйце развивается мирацидий (личинка материнской спороцисты) - первая личиночная стадия в жизненном цикле трематод. Мирацидий активно (самостоятельно внедряется) или пассивно (не выходит из яйца) попадает в моллюска. После заражения первого промежуточного хозяина (моллюска) в нем развиваются партеногенетические поколения - спороцисты и редии. Партениты в большинстве случаев представлены не одной, а несколькими генерациями, сменяющими друг друга. Завершающей стадией онтогенеза партеногенетических поколений является образование гермафродитного поколения трематод. В зародышевой полости дочерней партениты протекает морфогенез первой гермафродитной личинки - церкарии. Зрелые церкарии (похожи на микроскопических головастиков) выходят из моллюска и ведут активный образ жизни во внешней среде, как правило, водной. Затем они, либо инцистируются на субстрате и определяются как адолескарии
[у некоторых групп трематод (сем. Schistosomatidae) церкарии активно внедряются в тело дефинитивного хозяина], либо попадают (активным или пассивным путем) в организм второго промежуточного хозяина (беспозвоночные, рыбы, амфибии), где у большинства видов трематод проходят следующие стадии морфогенеза. Такая личинка обозначается как метацеркарий. Окончательное развитие осуществляется в дефинитивном хозяине - позвоночном животном и носит название маритогония. По ее завершении половозрелая особь гермафродитного поколения (марита) приступает к репродукции. Чередование двух способов полового размножения - партеногенетического и гермафродитного - указывает, что жизненный цикл трематод осуществляется по принципу гетерогонии.
Материнская спороциста.
Материнской спороцистой принято называть паразитирующую в организме моллюска-хозяина особь 1-го (материнского) партеногенетического поколения. В организме моллюска мирацидий претерпевает метаморфоз и становится материнской спороцистой. Превращение мирацидия в молодую материнскую спороцисту носит характер регрессивного метаморфоза. Полностью сформированная материнская спороциста обычно локализуется в гемоцеле или в мантии моллюска. Тело спороцисты представляет собой своеобразный «мешок», в котором заключены зародышевые клетки и возникающие в результате их дробления эмбрионы (зародышевые шары). Из них впоследствии развиваются особи следующего поколения партенит. Материнская спороциста имеет округлую, мешковидную или червеобразную форму, иногда бывает разветвленной (рис. 9). Она лишена кишечника и питается всей поверхностью своего тела. Центральная нервная система и органы чувств развиты слабо. Герминальный материал (полученный спороцистой от мирацидия) испытывает определенные преобразования, что определяет особенности организации и развития материнской спороцисты. Для одной группы трематод (сем. Fasciolidae, Paragonimidae, Echinostomatidae и др.) плодовитость материнской спороцисты ограничивается числом генеративных клеток, содержащихся в мирацидии. Следовательно, внутри материнской спороцисты число будущих редий или дочерних спороцист соответствует числу зародышевых шаров, находящихся в теле мирацидия. У этой группы трематод число особей следующего партеногенетического поколения, отрождаемых материнскими спороцистами, редко превышает 10. Другая группа трематод (сем. Alariidae, Stregeididae, Schistosomatidae и др.) обладает ясно выраженной способностью к умножению клеток генеративного ряда на стадии материнской спороцисты. Для них характерно формирование специального органа размножения - герминальной массы, которая функционирует на протяжении значительной части жизни спороцисты в организме первого промежуточного хозяина. Этот процесс называется полиэмбрионией и приводит к колоссальному увеличению числа зародышей, развивающихся в материнской спороцисте. В результате развития из зародышевых шаров в материнской спороцисте формируются особи второго (дочернего) партеногенетического поколения. Зрелые формы разрывают тело материнской спороцисты, и далее они мигрируют в печень моллюска.
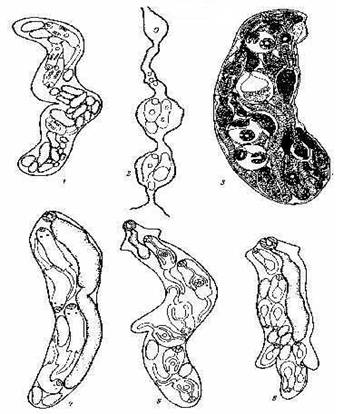
Рис.9. Партеногенетические поколения трематод: 1, 2 - материнские спороцисты (1 - мешковидная, 2- червеобразная); 3 - дочерняя спороциста; 4, 5, 6 - редии (4 - мешковидная, 5 - с локомоторными выростами и коротким кишечником (Fasciola hepatica), 6 - с локомоторными выростами и длинным кишечником).
Дочерние партеногенетические поколения
Дочернее партеногенетическое поколение может быть представлено двумя морфо-биологическими формами: редиями и спороцистами, которые между собой различаются как строением, так и биологией. Редии характерны обычно для более примитивных трематод (сем. Fasciolidae, Dicrocoeliidae, Echinostomatidae, Opisthorchidae, Paramphistomatidae и др.) (рис. 9). Они имеют вытянутое в длину цилиндрическое тело, на поверхности которого нередко располагаются конические локомоторные выросты. На переднем конце тела редий терминально открывается ротовое отверстие. Чуть позади субтерминально располагается родильная пора - небольшое отверстие, служащее для выхода отрождаемых особей. У редий имеется пищеварительная система, состоящая из глотки, пищевода и мешковидного кишечника. Имеется протонефридиальная выделительная система и железистый аппарат. Редии питаются активно, пожирая ткани печени моллюска. Основной объем тела редий занимает зародышевая полость, в которой протекают последовательные этапы развития особей следующего поколения. Размножение редий осуществляется партеногенетическим путем. В задней части редий находится небольшой компактный яичник, формирующий зародышевые клетки, которые дают начало эмбрионам следующего поколения - дочерним редиям или церкариям. Обычно продуктивность редий не очень велика, одновременно в ней могут находиться 2-3 дочерние редии или 6-10 сформированных церкарий. Как правило, вначале развивается несколько поколений редий, после этого редии начинают отрождать церкарий. За сутки из зараженного моллюска выходит в воду несколько десятков, реже сотен церкарий. Продолжительность жизни одной редии обычно не превышает 2-2,5 месяцев. Дочерние спороцисты характерны для более продвинутых в эволюционном плане групп трематод (сем. Alariidae, Stregeididae, Plagiorchidae и др.) (рис.9). Дочерние спороцисты - поколение гомологичное редиям. Они рассматриваются как недоразвитые редии, которые приобрели способность размножаться, еще не будучи до конца сформированными (Гинецинская, Добровольский, 1978). Дочерние спороцисты отличаются от редий значительно большей простотой строения. Локомоторные выросты отсутствуют, родильная пора, если имеется, располагается терминально. Пищеварительная система полностью редуцируется, а железистый аппарат по сравнению с редиями вторично упрощен. Форма дочерних спороцист может быть самой разнообразной: мешковидной, червеобразной, нитевидной, круглой и т. д. Дочерние спороцисты в молодом возрасте подвижны и имеют хорошо развитый кожно-мускульный мешок. С возрастом спороцисты становятся менее активными, а некоторые и вовсе утрачивают подвижность, буквально врастая в ткани печени моллюска, которую они пронизывают во всех направлениях. В отличие от материнских спороцист дочерние спороцисты имеют на переднем конце тела небольшое отверстие – «родильную пору», через которую выходят наружу отрождаемые спороцистой особи следующего поколения. Дочерние спороцисты способны отрождать как подобных себе дочерних спороцист, так и церкарий. Данный процесс называется полиэмбрионией и приводит к колоссальному увеличению числа зародышей. Подсчитано, что из одного зараженного моллюска в течение суток могут выходить десятки тысяч церкарий. Например, моллюск Littorina littorea, зараженный одним мирацидием Cryptocotyle lingua, за 7 лет наблюдений выделил несколько миллионов церкариев (Шульц, Гвоздев, 1970).
Гермафродитное поколение
Личиночные стадии гермафродитного поколения в ходе онтогенеза особей гермафродитного поколения трематод сменяются несколько последовательных стадий, каждая из которых приурочена к определенной среде обитания. Церкарий (рис. 10). Морфогенез этой первой личиночной стадии гермафродитного поколения протекает в зародышевой полости дочерней партениты и может завершаться уже во время ее миграции по телу первого промежуточного хозяина. По выходу из моллюска зрелый церкарий ведет активный образ жизни во внешней среде, как правило, водной. Внешне он напоминает микроскопического головастика, имеет овальное вытянутой формы тело и хвостовой придаток (хвост). Размеры тела колеблются от 0,1 до 0,8 мм. Хвост снабжен хорошо развитой мускулатурой. Строение хвоста варьирует у личинок различных видов трематод. Он может иметь вилообразную, плавниковообразную, булавообразную и другие формы, в т.ч. на хвосте могут находиться разнообразные придатки. У некоторых видов хвост может быть сильно укороченным или вовсе отсутствовать. Подобное строение хвоста обеспечивает церкариям разнообразные локомоторные функции: плавание и парение в толще воды, закрепление и ползание по субстрату и др. У церкариев уже сформированы сходные с маритами органы: присоски, пищеварительная система, органы выделения (от марит отличаются лишь меньшим числом мерцательных клеток), хорошо развита нервная система. Органы чувств представлены многочисленными сенсиллами. Наибольшее число сенсилл сосредоточено на переднем конце тела, где располагается мозговой ганглий. У многих видов имеются глаза, которые топографически и функционально связаны с мозговым ганглием (рис. 10). В теле церкариев находятся многочисленные железистые клетки, выполняющие различные функции. Выявлены три основных типа желез: цистогенные, слизистые и железы проникновения. У личинок, инцистирующихся во внешней среде, хорошо развиты цистогенные железы, которые располагаются в самых поверхностных слоях паренхимы. Секрет этих желез идет на построение толстой оболочки цисты при инцистировании следующей стадии развития (адолескария). Слизистые железы располагаются вдоль средней линии тела. Их секрет окутывает тело церкария и, с одной стороны, защищает личинку от вредного воздействия пищеварительных ферментов моллюска-хозяина, с другой - способствует прикреплению церкария к телу второго промежуточного хозяина. У тех церкариев, которые активно внедряются в тело второго промежуточного хозяина и инцистируются в нем, хорошо развиты железы проникновения. Как правило, эти железы располагаются вокруг брюшной присоски, и их протоки открываются на переднем конце тела. С помощью выделяемого ими секрета церкарии легко внедряются через покровные ткани животных-хозяев. Вышедшие из моллюска церкарии не питаются. Они живут за счет запасов гликогена, который накапливается в их теле в процессе развития. Особенно много гликогена сосредоточено в хвосте, что связано с активной локомоторной функцией этого органа. В среднем церкарии живут 24-48 часов. Личинки не «сумевшие» найти хозяина элиминируются.
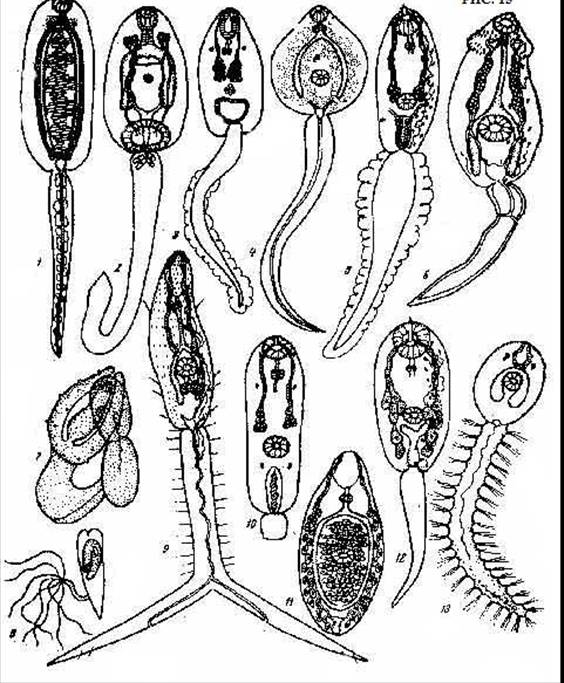
Рис. 10. Различные типы церкариев трематод: 1 - одноприсосковые, 2 - заднеприсосковые, 3 - плавниково-гребенчатые (Opisthorchis felineus), 4 - невооруженные (Fasciola hepatica), 5 - булавохвостые, 6 - эхиностоматидные, 7 - мешкохвостые, 8 - котилюрные, 9 - вилохвостые, 10 - короткохвостые, 11 - бесхвостые, 12 - стилетные, 13 - щетинкохвостые (по Шульц, Гвоздеву, 1970).
На данном этапе развития основные биологические функции церкариев направлены на активный поиск соответствующего «субстрата» (субстраты внешней среды или тело второго промежуточного хозяина) и последующую цистогонию (инцистирование на субстрате или в теле хозяина). В результате этих биологических «событий» формируется следующая стадия гермафродитного поколения. Церкарии обладают широким набором адаптаций, направленных на успешный поиск субстрата. Важную роль в этом процессе играют таксисы
(фото-, гео- и хемотаксисы), благодаря которым церкарии находят необходимый субстрат. Например, описторхидные (сем. Opisthorchidae) церкарии парят в толще воды в виде скоплений (облаков) и «ждут» появления второго промежуточного хозяина - карповых рыб. В случае появления хозяина церкарии атакуют рыбу, закрепляются на поверхности ее тела, отбрасывают хвост, а затем проникают при помощи секрета желез проникновения в мышечную ткань. Также достаточно выраженным хемотаксисом обладают представители сем. Stregiidae, церкарии которых внедряются в тело пиявок определенного вида, реагируя на выделяемую ими слизь. Следующая фаза в биологии церкариев определяется как цистогония, т.е. процесс превращения церкария в неподвижную инцистированную (заключенную в цисту) личинку снаружи или внутри «субстрата». Выделяются три основные формы проявления цистогонии. 1.У некоторых трематод церкарии не покидают моллюска и в нем же и инцистируются. По сути, эти особи моллюсков превращаются во второго промежуточного хозяина. 2. Церкарии инцистируются на субстрате в водной среде: непосредственно в воде (как правило, на поверхностной пленке), на растениях, на раковинах моллюсков, на панцире ракообразных и т.д. 3. Церкарии инцистируются после проникновения в организм второго промежуточного хозяина. В процессе цистогонии церкарии одеваются плотной многослойной оболочкой (цистой) за счет секрета цистогенных желез. Биологическое значение цист сводится к защите личинки от неблагоприятных воздействий внешней среды или иммунологических реакций хозяина. Личинка, заключенная в цисту, превращается в следующую фазу развития - метацеркарий. Метацеркарий (рис. 11). Данная фаза возникла последней в ходе эволюции жизненных циклов трематод. Это обстоятельство, как считают современные исследователи, и определило то удивительное многообразие метацеркарий, которое обнаруживается в различных группах трематод (Галактионов, Добровольский, 1998). В жизненных циклах ряда трематод (сем. Fasciolidae, Notocotylidae, Echinostomatidae и др.) присутствует фаза адолескария, т.е. личинки, инцистирующейся во внешней среде (рис. 11). В процессе инцистирования на субстрате церкарий отбрасывает хвост, а из тегумента изливается наружу цистогенный секрет. Цисты адолескарий, как правило, многослойны, обладают большой механической прочностью, не проницаемы для воды и растворенных в ней химических веществ. Отличительными особенностями адолескария (в сравнении с церкарием) является отсутствие хвоста и цистогенных желез. Это свободная покоящаяся стадия, продолжительность жизни которой обусловлена запасами гликогена. В более продвинутых в эволюционном плане группах трематод (это огромное число трематод, включая сем. Dicrocoeliidae, Plagiorchidae, Alariidae, Stregiidae и др.) в жизненном цикле появляется второй промежуточный хозяин. В момент внедрения в промежуточного хозяина церкарий отбрасывает хвост и, добравшись до места обычной своей локализации (мышцы, полость тела, подкожная клетчатка, глаза и т. д.), инцистируется, превращаясь в метацеркария. Циста (в отличие от адолескариев) в той или иной степени редуцируется и становится более тонкостенной, состоит, как правило, из 2-х слоев (рис. 11). Внутри цисты метацеркарий претерпевает развитие (морфогенез), в его паренхиме откладываются запасы гликогена. Во время морфогенеза метацеркарии растут, увеличиваются значительно в размерах. Этому процессу предшествует резорбция некоторых органов, характерных 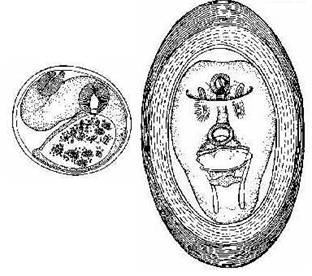 для церкариев. У метацеркариев увеличивается число мерцательных клеток, формируются новые железы, развиваются половые железы, иногда появляется даже сумка цирруса и зачаток матки. В частности, метацеркарии трематод отр. Stregiidida претерпевают настоящий метаморфоз, напоминающий полный метаморфоз насекомых. У таких видов сначала разрушаются все органы, присущие церкариям, после чего происходит формирование новых органов, свойственных метацеркариям.
для церкариев. У метацеркариев увеличивается число мерцательных клеток, формируются новые железы, развиваются половые железы, иногда появляется даже сумка цирруса и зачаток матки. В частности, метацеркарии трематод отр. Stregiidida претерпевают настоящий метаморфоз, напоминающий полный метаморфоз насекомых. У таких видов сначала разрушаются все органы, присущие церкариям, после чего происходит формирование новых органов, свойственных метацеркариям.
Рис. 11. Метацеркарии: а - Opisthorshis felineus, инцистированный в мышцах карповых рыб (Шульц, Гвоздев, 1970); б - Strigea sphaerula, из ткани моллюска, снабженная толстой капсулой (по Гинецинской, 1968).
Следует отметить, что развивающийся метацеркарий питается извне, т.е. из тканей животного - хозяина. Так же установлено, что наружные оболочки вокруг инцистированных личинок образуются тканями хозяина, т.е. формируется капсула. Следовательно, в организме промежуточного хозяина циста с метацеркарием помещается внутри капсулы. После завершения морфогенеза метацеркарии становятся инвазионными и способны заражать дефинитивного хозяина. У некоторых групп трематод, а это представители отр. Stregiidida (роды Alaria и Strigea), в ходе онтогенеза особей гермафродитного поколения появляются дополнительные этапы. В жизненный цикл между фазами церкарий и метацеркарий вклинивается дополнительная фаза развития - мезоцеркарий. Хозяин, в котором формируется мезоцеркарий, называется «вставочным», и он встраивается соответственно между первым и вторым промежуточными хозяевами. В качестве вставочного хозяина у этой группы трематод зарегистрированы бесхвостые амфибии. Мезоцеркарий лишен хвоста, который отбрасывается при внедрении, и значительно увеличивается в размерах по сравнению с церкарием (Судариков, 1984). В то же время вооружение, пищеварительная система, половой зачаток мезоцеркария остаются такими же, как и у церкария. Сохраняются и железы проникновения, объем которых сильно возрастает. Мезоцеркарий неинвазионен для окончательного хозяина и должен пройти в своем развитии фазу метацеркария, которая характеризуется диаметрально противоположными морфогенетическими тенденциями. Появление в жизненном цикле трематод мезоцеркария как самостоятельной личинки и переход от триксенного типа развития к тетраксенному - явление прогрессивное. Оно способствовало укреплению связи трематод, развивающихся с участием водных организмов, с наземными животными, не имеющих прочных экологических связей с водной средой.
Жизненные циклы трематод. Как следует из предыдущего раздела, для трематод характерны сложные жизненные циклы, связанные с чередованием поколений и сменой хозяев. Чередование поколений (гермафродитного и партеногенетического) в жизненных циклах трематод определяется как гетерогония. В некоторых руководствах до сих пор партеногенетическое размножение неправильно считается бесполым и отсюда за жизненным циклом трематод неправильно признается метагенез (т.е. чередование полового и бесполого поколений), что не соответствует действительности. Полный и наиболее сложный цикл развития трематод происходит при участии трех хозяев: окончательного (дефинитивного) и двух промежуточных (первого и второго). Первым промежуточным хозяином всегда служит моллюск. Вторым промежуточным хозяином могут являться различные беспозвоночные (моллюски, полихеты, ракообразные, насекомые) и позвоночные (рыбы, амфибии, рептилии, птицы, млекопитающие). В реализацию жизненного цикла у небольшой группы трематод между первым и вторым промежуточными хозяевами могут вклиниваться вставочные хозяева (высшие и низшие позвоночные животные). В некоторых, экологически детерминированных, случаях в жизненный цикл отдельных групп трематод могут включаться резервуарные хозяева. Резервуарными хозяевами определяют таких животных, которые, обладая возможностью накапливать в себе инвазионных личинок гельминта, способствуют передаче их дефинитивному хозяину. Однако резервуарные хозяева не являются обязательным звеном в ходе жизненного цикла. Особенно часто резервуарные хозяева как экологически детерминированное звено отмечается в системах хищник - жертва.
СИСТЕМАТИКА ТРЕМАТОД
Классификация трематод началась с К.Линнея (1758), признававшего один род Fasciola с двумя видами. В последующем были предприняты попытки систематики трематод на основе морфологии половозрелых трематод, строения экскреторной системы и внешних признаков церкариев и биологии паразитов. Значительный вклад в систематику трематод внесли гельминтологи К.И.Скрябин и Р.С.Шульц. Ниже приводится систематика трематод, принятая в большинстве стран.
КЛАСС Trematoda (Rudolphi, 1808).
1.ПОДКЛАСС Bucephalidae – ротовое отверстие на брюшной стороне тела, около его середине, лишено ротовой присоски. Кишечник простой, мешковидный, короткий. На переднем конце имеется мышечная присоска или хоботок со щупальцами. Подкласс включает один отряд и одно семейство. Паразиты карповых, окуневых, лососевых, тресковых (налим) рыб и амфибий.
ОТРЯД Bucephalida
Сем. Bucephalidea (Poche, 1907).
Семейство объединяет несколько родов и более 100 видов.
2.ПОДКЛАСС Prosostomidea (Skrjabin et Gusochanskaja, 1962) – органы прикрепления представлены различно развитыми ротовой и брюшной присосками, одна из них или обе могут отсутствовать. Развитие сложное: с чередованием поколений и сменой хозяев. Подкласс состоит из двух отрядов.
1. ОТРЯД Fasciolida (Skrjabin et Guschanskaja, 1962), включает 12 подотрядов:
а) ПОДОТРЯД Schistosomatata (Skrjabin et Schulz, 1937)
- раздельнополые трематоды, присоски развиты слабо или вовсе отсутствуют; яйца обычно с шипиками. Паразитируют в кровеносной системе птиц и млекопитающих;
б) ПОДОТРЯД Sanquinicolata (Skrjabin et Schulz,1937) - гермафродитные трематоды, паразиты крови рыб и рептилий. Присоски имеются или отсутствуют, фаринкса нет, матка короткая. Включает сем. Spirorchidae и Sanquinicolidae. Последние мелкие трематоды (2 мм длины), матка у них отсутствует, в оотипе имеется только одно яйцо, иногда имеется укороченная матка. В рыбах пресных водоемов РФ встречаются восемь видов из рода Sanquinicola, но наиболее патогенен S.inermis;
в) ПОДОТРЯД Pronocephalata (Skrjabin,1955) – брюшная присоска и фаринкс отсутствуют, половые железы расположены в задней части тела. Паразиты пищеварительного тракта и органов дыхания позвоночных. Наиболее часто встречаются виды из семейств:
сем. Notocotylidae (Luhe, 1909) – мелкие и средней величины трематоды (3-5 мм длины), тело в задней части густо усеяно мельчайшими шипиками. Ротовая присоска простая, пищевод короткий, фаринкс отсутствует. Семейство содержит 9 родов; у домашних водоплавающих птиц паразитируют представители родов Notocotylus (N. аttenuatus, N. chionis), Catatropis (С. verrucosa).
сем. Pronocephalidae (Looss, 1902) – мелкие и средние трематоды, передний конец снабжен воротничком, на заднем – два выступа; яйца многочисленные, с филаментами. Паразитируют в кишечнике морских и пресноводных черепах;
г) ПОДОТРЯД Cyclocoeliata (La Rue, 1957) – крупных и мелких размеров трематоды, кутикула без шипов, ротовая присоска отсутствует, брюшная присоска рудиментарна. Зрелые яйца в матке содержат мирацидиев, снабженных пигментным глазком; мирацидии могут выходить из скорлупы еще внутри матки. Паразитируют в полости тела, носовых и глазных полостях, в воздухоносных мешках приводных птиц, преимущественно диких (Tracheophilus sisowi);
д) ПОДОТРЯД Echinostomatata (Szidat, 1936) - имеют вооруженный или невооруженный адоральный диск (воротничок):
сем. Echinostomatidae (Dietz, 1909) – есть адоральный диск, снабженный шипиками. Кутикула в передней части имеет мелкие шипики, брюшная присоска крупная. Паразитируют в кишечнике, желчных ходах и фабрициевой сумке птиц. Это семейство содержит 40 родов, объединяющих около 300 видов. Наиболее богаты видами роды Echinostoma (E.revolutum, E.miyagawai, E.paraulum), Echinoparyphium (E. recurvatum), Echinochasmus (E.beleocephalus).
Сем. Cotylotretidae (Skrjabin at Baschkirova, 1956) – адоральный диск отсутствует, головной конец лишен шипов. Паразитируют у птиц, грызунов и ластоногих.
е) ПОДОТРЯД Heterophyata (Morosow, 1955) - мелкие и средней величины трематоды, снабжены ротовой и брюшной присосками. Церкарии лишены присоски, метацеркарии – в рыбах или ракообразных, половозрелые паразиты – в пищеварительном тракте, желчных ходах печени, желчном пузыре млекопитающих.
Сем. Heterophyidae (Odhner, 1914) – мелкие трематоды (2,5 мм), кутикула покрыта шипами. Развиваются с участием двух промежуточных хозяев (моллюски и рыбы). Паразитируют в кишечнике человека.
Сем. Galactosomidae (Morosow, 1950) – 3–3,5 мм длины, брюшная присоска маленькая. Паразитируют в кишечнике птиц и млекопитающих. Метацеркарии локализуются в чешуе, жабрах, плавниках и других органах рыб.
Сем. Opisthorchidae (Braun, 1901) – средней величины трематоды; присоски сближены; брюшная присоска слабо развита. Яйца мелкие, коричневого цвета. Семейство объединяет более 20 родов (80 видов). Наибольшее значение имеют роды: Opisthorchis (O.felineus), Clonorchis (C.sinensis) – паразиты печени у человека.
ж) ПОДОТРЯД Azygiata (La Rue, 1957) - трематоды имеют ротовую и брюшную присоски, мышечный фаринкс. Половозрелые трематоды паразитируют в кишечнике рыб (окуней, щук);
з) ПОДОТРЯД Allocreadiata (Skrjabin, Petrow et Kovae 1958) - овальные или удлиненные трематоды с хорошо развитыми присосками. Яйца крупные, с крышечками. Паразиты пищеварительного тракта судаков, окуней, щук, лососей, хариусов, налимов (сем. Bunoderidae, Nicoll, 1914 – Bunodera Euciopercae, Crepidostomum farionis), пресноводных и морских рыб (сем. Opecoelidae, Ozaki, 1925);
и) ПОДОТРЯД Fasciolata (Skrjabin et Schulz, 1935) – трематоды с очень разнообразной формой тела, размером от 1 мм до нескольких см. К этому отряду относятся более 40 семейств. Наибольшее значение имеют семейства:
Сем. Fasciolidae (Railliet, 1895) – крупные трематоды с близко расположенными ротовой и брюшной присосками, семяприемник отсутствует. Семейство включает до 10 видов пяти родов, паразитирующих в желчных ходах печени (реже в кишечнике) наземных млекопитающих (копытных), а также человека. Широко распространены и наиболее патогенны следующие виды: Fasciola hepatica, F.gigantica (в РФ), F.californisa, F.halli в (США). Развиваются с одним промежуточным хозяином – моллюсками. Адолескарии инцистируются на водных и прибрежных растениях или на поверхности воды.
Сем. Dicrocoeliidae (Odhner, 1911) – удлиненные либо круглые трематоды. Присоски сближены, лежат в передней части тела, фаринкс развит хорошо, имеется пищевод. Хорошо развита половая бурса, содержит невооруженный циррус, семенной пузырь.
Вид Dicrocoelium lanceatum – ланцетовидная двуустка – паразит желчных протоков и желчного пузыря копытных, зайцеобразных и грызунов. Развитие происходит со сменой двух промежуточных хозяев – наземных моллюсков и насекомых (муравьев).
Вид Eurytrema pancreaticum – паразитирует в протоках поджелудочной железы у крупного и мелкого рогатого скота.
Сем. Plagiorchidae (Luhe, 1901) – мелкие трематоды, кутикула их покрыта шипиками. Половое отверстие внутри брюшной присоски, семенники – позади яичника, половая бурса длинная, заходит за уровень брюшной присоски, внутри содержит семенной пузырек. Циррус короткий или длинный. Матка располагается позади брюшной присоски, проходит между семенниками, достигая заднего конца тела. Яйца мелкие, церкарии имеют стилет. Семейство включает много родов, но наибольшее значение имеет род Plagiorchis, вид P.orcuatus – паразит яйцевода и фабрициевой сумки домашних кур. P. maculosus – диких птиц, P.cutamiatis – ондатры.
Сем. Prostogonimidae (Nicoll, 1924) – мелких и средних размеров трематоды, с суженным передним и расширенным задним концом тела. Кутикула покрыта шипиками. Паразиты яйцевода, фабрициевой сумки птиц. Паразиты развиваются с двумя промежуточными хозяевами – моллюсками и насекомыми. Типичный род - Prosthogonimus; наибольшее ветеринарное значение имеют P.ovatus P. cuneatus P.macrorchis;
к) ПОДОТРЯД Paramphistomatata (Szidat, 1936), (Skrjabin
et Schulz, 1937) - имеет сдвинутую к заднему концу тела брюшную присоску, кишечные стволы мощно развиты. Развиваются при участии одного промежуточного хозяина. Церкарии во внешней среде превращаются в адолескариев. Паразиты рыб, амфибий, птиц и млекопитающих. Подотряд включает десять семейств, наибольшее ветеринарное значение имеют семейства:
Сем. Paramphistomatidae (Fischceder, 1901) – тело коническое, цилиндрическое (2-12 мм), кутикула без шипиков. Задняя присоска лежит апикально, реже вентрально. Половая бурса отсутствует. Паразитирует в желудке, рубце и желчных ходах копытных млекопитающих. В это семейство входят девять родов, объединяющих около 60 видов. У копытных в РФ зарегистрировано несколько родов Paramphistomum, Calicophoron, Liorchis.
Сем.Gastrothylacidae (Stiles et Goldberger, 1910) - распространены в странах с жарким климатом, описано 20 видов, в нашей стране встречаются 2 вида: Gastrothylaх crymenifer (1847) и Fischoederius elongatus (1883).
2. ОТРЯД Strigeidida (La Rue, 1926), (Sudaricov, 1959) – имеют позади брюшной присоски орган Брандеса – сложный секреторно - фиксаторный аппарат; половое отверстие находится на заднем конце тела. Паразитируют в кишечнике рептилий, птиц и млекопитающих. Первыми промежуточными хозяевами являются брюхоногие моллюски, вторыми – беспозвоночные (моллюски, пиявки, олигохеты) и позвоночные всех классов. Отряд включает два подотряда Strigeata, Cyatocotylata, из которых наибольшее ветеринарное значение имеет первый подотряд, включающий восемь семейств.
Сем. Strigeidae (Railliet, 1919) – чашевидный передний сегмент отделен от заднего сужением или перетяжкой. Половое отверстие вблизи заднего конца тела. Семенники позади яичника, матка короткая (содержит мало яиц), петли ее простираются не дальше межсегментной границы. Мариты паразитируют в кишечнике птиц. Развиваются с двумя промежуточными хозяевами. Вторым промежуточным хозяином являются моллюски, пиявки, рыбы, рептилии, птицы. Семейство объединяет около 110 видов, наиболее распространены виды из родов Strigea, Cotilurus, Apatemon.
Сем. Diplostomatidae (Poirier, 1886) – рожковидные и лопатовидные трематоды, орган Брандеса в виде округлой присоски. На передней крае тела имеются псевдоприсоски. Матка и семяизвергательный канал, сливаются в непарный гермафродитный канал, открывающийся в полость полового атриума. Семейство включает 23 рода, наибольшее ветеринарное значение имеют представителей родов Diplostomum, Neodiplostomum, Postodiplostomum, Tylodelphue. Мариты паразитируют в кишечнике приводных птиц, первый промежуточный хозяин – пресноводные моллюски, второй – многие пресноводные рыбы, в хрусталике которых формируются метарцеркарии.
Сем. Alariidae-мелкие трематоды (2,4-4,4), вокруг ротовой присоски имеются ушковидные выросты. Мариты - Alaria alata паразитируют в тонком кишечнике плотоядных. Развиваются с участием двух промежуточных хозяев (моллюски, амфибии).
СИСТЕМАТИКА ТРЕМАТОД
Тип PLATHELMINTHES - плоские гельминты. Класс TREMATODA






