К настоящему моменту известно, что вариации поверхностных антигенов могут быть реализованы с помощью двух механизмов. Первый - использование сигнальных путей для получения информации о микроокружении клетки и ответ путем экспрессии продуктов генов вирулентности, необходимых для выживания в организме хозяина [54]. Другой механизм - спонтанная и случайная генерация клеточных популяциий с различными антигенными фенотипами, или "гетеротипов", которые могут выдержать атаку макроорганизма, способную уничтожить исходную популяцию. Частота возникновения таких антигенных вариаций очень высока (10-4-10-2 на клетку на генерацию) по сравнению со спонтанными мутациями (10-6-10-8)[52]. Данные сиквенирования геномов M.pneumoniae и M.genitalium показывают, что их геномы не содержат регуляторных генов, кодирующих сигнальные белки, ответственные за определение условий микроокружения. Также показано очень маленькое количество белков-шаперонов, ответственных за белковую секрецию, отсутствие сигнальной пептидазы I типа [28,26]. Эти данные свидетельствуют о низкой вероятности для Mollicutes осуществлять антигенную вариабельность по первому пути.
Несмотря на очень маленький размер генома, число генов Mollicutes, вовлеченных в создание антигенных структур у них очень высоко. Общим мотивом у патогенных микроорганизмов и паразитов является наличие множественных вариабельных генов, объединенных в семейства. Переключение каждого гена во включенное или выключенное состояние происходит с высокой скоростью, к тому-же каждый ген способен производить продукты разной длины. Таким образом осуществляется синтез большого числа различных поверхностных антигенных структур. [52] Стоит отметить наличие семейств генов у M.gallisepticum (pMGA семейство), M.bovis (Vsp семейство), M.hyorhinis (Vlp семейство), M.pulmonis (Vsa семейство) и другие.
Генетические механизмы реализации вариабельности поверхностных антигенов можно разделить на три категории.
Вариации, обусловленные наличием гомополимерных повторов. Особенностью данного типа генетических вариаций является наличие небольших регионов, содержащих повторяющиеся основания или олигонуклеотидные повторы. Такие "горячие точки" являются отличной мишенью для частых вставок и делеций нуклеотидов, таким образом позволяя включать или выключать экспрессию генов.
Например, промотор Vlp генов у M.hyorhinis (рис 1) содержит последовательность poly(A) непосредственно перед ТАТААТ боксом, правее региона -35. Этот участок длиной в 17 или 18 оснований позволяет гену нормально экспрессироваться, тогда как увеличение его длины до 20 оснований выключает экспрессию гена [21,73].

Рис. 1 Строение генов семейства Vlp.
Подобным образом функционируют семейства генов Vaa M.hominis (рис. 2) и P78 M.fermentas (рис. 3) [75,68].
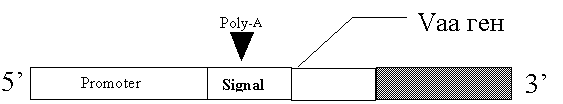
Рис. 2 Строение генов семейства Vaa. 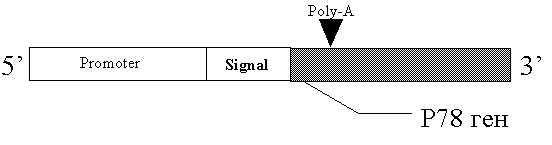
Рис. 3 Строение генов семейства P78.
У M.gallisepticum в качестве олигонуклеотидного повтора представлена последовательность (GAA)n,где n меняется от 10 до 16 (рис 4) [10].
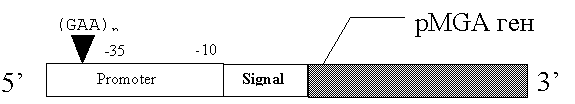
Рис. 4 Строение генов семейства pMGA.
Вариации, обусловленные наличием хромосомных перестановок. Инверсии ДНК, генная конверсия, дупликации или делеции тандемных гомологичных блоков ДНК, перемещение транспозонов также являются широко распространенным механизмом антигенной гиперизменчивости. Полагают, что большинство этих перестановок спонтанны и случайны, однако не исключается возможность и гомологичной рекомбинации, требующей для своей реализации активности RecA [25,67].
Например показано, что в формировании вариабельного поверхностного липопротеина M.bovis учавствуют хромосомные перестановки [39,38]. Механизм этих перестановок представлен на рисунке 5:
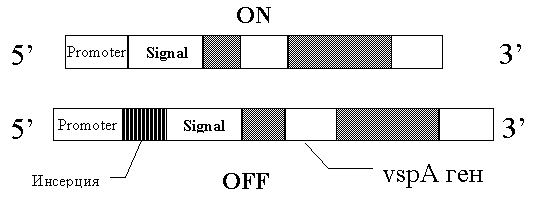
Рис. 5 Механизм формирования вариабельного поверхностного липопротеина: при инсерции нуклеотидной последовательности между промотером и участком ДНК, ответственным за синтез сигнального пептида, экспрессия антигена выключается
Другим примером, иллюстрирующим влияние хромосомных перестановок на экспрессию поверхностных антигенных структур, является семейство V-1 антигенов M.pulmonis [14,15,63]. Была найдена корреляция между изолятами с разными вариантами экспрессии V-1 и способностью этих изолятов к гемадсорбции (HA фенотип). Анализ нуклеотидной последовательности гена V-1 двух фенотипов (vsa HA+ и vsa HA-) показал, что эти фенотипы возникают в результате хромосомной инверсии. У "молчащего" гена vsa отсутствует 5' регион, содержащий промотер и сайт связывания рибосомы, которые присутствуют в экспрессирующемся гене (рис 6):
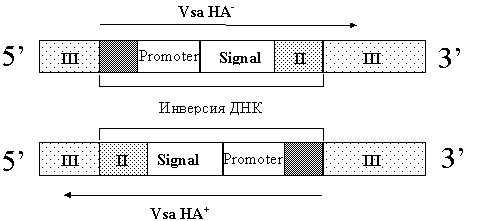
Рис. 6 Механизм переключения между HA- и НA+ фенотипами: при инверсии региона, содержащего промотер и последовательность сигнального пептида возникает переключение экспресии гена V-1.
Вариации, обусловленные наличием повторяющихся доменов (вариабельность размера). Среди молекул, вовлеченных во взаимодействие микроорганизм-хозяин найдено много поверхностных антигенов с тандемными повторяющимися доменами. Роль этих доменов напрямую связана со связыванием лиганда и образованием антигенных вариаций [24,32,34]. Наличие множества тандемно расположенных доменов в генах, кодирующих антигены, является удобным инструментом для образования антигенов различной длины. Молекулярным механизмом, обеспечивающим делеции и вставки целых доменов, является гомологичная рекомбинация [36]. Не удивительно, что и микоплазмы обладают большим количеством подобных антигенов. Это гены Vlp M.hyorhinis [59,60], Vsp M.bovis [12,39,58], МВ антиген U.urealyticum [77], Vaa адгезин и Lmp1 антиген M.hominis [74], P30 и P1 M.pneumoniae [22], V-1 антиген M.pulmonis [63].
В последнее время повышенный интерес у исследователей вызывает так называемый Multiple banded antigene или МВ-антиген уреаплазм. Показано, что его длина варьрут у различных клинических изолятов [70, 71, 76]. Этот факт имеет важное значение, так как вариабельность длины у подобного антигена Mycoplasma pulmonis является единственным отличием между вирулентными и невирулентными штаммами [71].
Фенотипическая характеристика МВ-антигена показывает, что он: а) видоспецифичен; б) содержит как сероварспецифичные, так и перекрестно реагирующие эпитопы; в) синтезируется как in vivo так и in vitro; г) подвергается высокому уровню изменчивости размера in vitro; д) присутствует в инвазивных изолятах U.urealyticum; е) является доминирущим антигеном, узнаваемым при инфицировании человека U.urealyticum [70].
Анализ ДНК последовательности гена МВ позволяет судить о структуре МВ-антигена (рис 7)
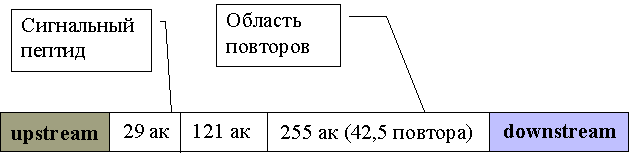
Рис. 7 Структура гена МВ.
Он представляет собой пептид с теоретической массой 42.863, содержащий на N-конце сигнальный пептид, а на С-конце повторяющуюся последовательность из 6 аминокислот (Glu-Gln-Pro-Ala-Gly). Полагают, что МВ-антиген своим N-концом прикреплен к мембране, выставляя С-конец во внешнюю среду [70].





