Типы ионных каналов и их относительное содержание в мембранах нервных, сердечных и гладкомышечных клеток неодинаково. Свойства ионных каналов в мембране каждой клетки изменялись в процессе эволюции, чтобы соответствовать ее функциональным потребностям. Сборка ионного канала - очень сложный процесс, поскольку при взаимодействии α-субъединиц и регуляторных р-субъединиц разных типов может формироваться большое количество подтипов даже одного вида канала. Более того, при альтернативном сплайсинге матричной РНК иногда образуется много изоформ одного и того же канала. Несмотря на эту сложность, экспрессия ионного канала, форма потенциала действия и функция клеток в различных типах тканей взаимосвязаны (рис.10).
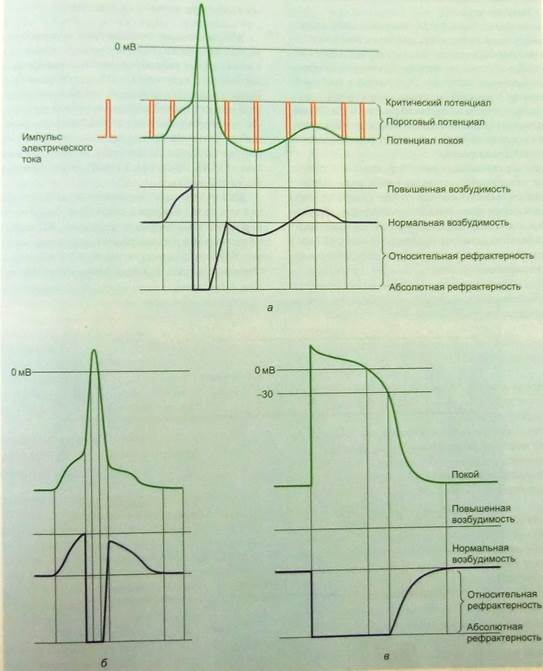
Рис. 10 Типы потенциалов действия и соотношение возбудимости с фазами ПД в нервном волокне (а), мышечном поперечнополосатом волокне (б) и в клетке рабочего миокарда желудочков сердца (в).
Потенциал действия нервной клетки. Нервные клетки должны возбуждаться быстро и многократно, чтобы передавать по нервной системе электрические импульсы. Передавать информацию другим нервным клеткам или рабочим органам – это основная функция потенциала действия нейронов. Их потенциал действия отражает эту функциональную потребность. Ему свойственны быстрые колебания и короткая продолжительность. Продолжительность спайка от 0,4 до 2 мс. Быстраая активация Na+- каналов обеспечивает крутой подъем кривой потенциала действия. Их почти мгновенная инактивация, сопровождающаяся активацией К+- каналов, обусловливает быстрое развитие реполяризации. При развитии потенциала действия активируются также потенциалзависимые Ca2+- каналы нейронного типа (N- типа), и происходящий при этом вход Ca2+ через пресинаптическую мембрану запускает выделение нейромедиатора. Ca2+- каналы N– типа быстро инактивируются, что позволяет следующему потенциалу действия прийти в тот момент, когда медиатор уже не выделяется. Тем самым достигается дискретность передачи информации.
Потенциал действия волокна поперечнополосатой мышцы по механизму образования и виду практически не отличается от потенциала действия нервной клетки. У него немного меньше крутизна нарастания фазы деполяризации и шире спайк. Это связано с функцией потенциала действия в скелетных мышцах. Основная задача этого потенциала действия, быстро возникнув, быстро распространиться внутрь мышечного волокна и вызвать выход ионов кальция из саркоплазматического ретикулума для мышечного сокращения.
В клетках миокарда желудочков сердца мембранный потенциал составляет -90мВ. Их потенциал действия в несколько сотен раз длительнее потенциала действия нейронов (рис.10). Большая продолжительность потенциала действия кардиомиоцита рабочего миокарда желудочков отражает функциональное назначение этих мышечных клеток, которое заключается в сокращении и расслаблении желудочков сердца с относительно небольшой частотой от 60 до 90 сокращений в минуту. Благодаря длительному потенциалу действия, увеличена и фаза абсолютной рефрактерности, которая, приблизительно, равна длительности систолы, что не дает миокарду возможности образовывать тетанические сокращения. Сердечная мышца работает в норме только в режиме одиночного мышечного сокращения, что отличает эту мышцу от скелетной мышцы.
Скелетная мышца из-за короткого потенциала действия способна работать и в режиме одиночного мышечного сокращения, и в режиме тетанических сокращений. Стоит отметить, что вклад потенциалзависимых Na+- и К+- каналов в фазы быстрой деполяризации и реполяризации потенциала действия кардиомиоцита аналогичен их роли в процессе возбуждения нервных клеток. На плазматической мембране кардиомиоцитов экспрессируется другой тип потенциалзависимых Ca2+- каналов, отличный от обнаруживаемого в нервных клетках. Деполяризация кардиомиоцитов вызывает активацию долговременных (L-тип) потенциалзависимых Ca2+- каналов, которые формируют продолжительный вход Ca2+ из внутриклеточного депо, обеспечивая более длительную деполяризацию и длительную фазу плато потенциала действия. Кроме того, этот приток Ca2+, сопряженный с выходом его из внутриклеточных депо, обеспечивает активирующую концентрацию Ca2+, необходимую для интенсивного сокращения желудочков сердца. Длительность плато в потенциале действия кардиомиоцита желудочков сердца является одним из механизмов регуляции силы сокращения сердца.
Гладкомышечные клетки. Гладкомышечные клетки представлены в различных тканях, в том числе в кровеносных сосудах, мочевом пузыре, матке и желудочно-кишечном тракте. Электрические свойства этих клеток в разных типах тканей значительно различаются. Основные отличия электрических свойств этих клеток от нервных, кардиомиоцитов и волокон скелетной мышцы заключаются в низком содержании Na+- каналов в плазматической мембране и более высоком мембранном потенциале покоя (от-45 до -60мВ). При таком мембранном потенциале потенциалзависимые Na+- каналы (если они есть) в значительной степени инактивированы и не могут участвовать в возбуждении клетки. Таким образом, основную роль в возбуждении гладкомышечных клеток играют потенциал-зависимые Ca 2+ - каналы L – типа. Вход Ca2+- в клетку через кальциевые каналы обеспечивает возникновение потенциала действия и достаточно высокую для мышечного сокращения внутриклеточную концентрацию кальция. Конфигурация этих потенциалов действия может быть растянутой или заостренной. Кроме того, повышение концентрации ионов кальция в большинстве гладкомышечных клеток происходит благодаря не потенциалу действия, а ступенчатым изменениям мембранного потенциала покоя. Характер возбуждения различных видов гладкомышечных клеток отражает их функцию в организме. Некоторые гладкомышечные клетки желудочно-кишечного тракта, участвующие в перестальтике, имеют ритмичные Ca2+- зависимые потенциалы действия. Гладкомышечным клеткам сосудов для поддержания артериального тонуса и предотвращения быстрых перепадов кровяного давления требуется постоянный приток Ca2+. Поэтому в этих клетках поддерживается относительно постоянный уровень потенциала покоя, сопряженного с потенциал-зависимым притоком Ca2+.





