

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...

Наброски и зарисовки растений, плодов, цветов: Освоить конструктивное построение структуры дерева через зарисовки отдельных деревьев, группы деревьев...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Наброски и зарисовки растений, плодов, цветов: Освоить конструктивное построение структуры дерева через зарисовки отдельных деревьев, группы деревьев...
Топ:
Теоретическая значимость работы: Описание теоретической значимости (ценности) результатов исследования должно присутствовать во введении...
Процедура выполнения команд. Рабочий цикл процессора: Функционирование процессора в основном состоит из повторяющихся рабочих циклов, каждый из которых соответствует...
Марксистская теория происхождения государства: По мнению Маркса и Энгельса, в основе развития общества, происходящих в нем изменений лежит...
Интересное:
Наиболее распространенные виды рака: Раковая опухоль — это самостоятельное новообразование, которое может возникнуть и от повышенного давления...
Лечение прогрессирующих форм рака: Одним из наиболее важных достижений экспериментальной химиотерапии опухолей, начатой в 60-х и реализованной в 70-х годах, является...
Распространение рака на другие отдаленные от желудка органы: Характерных симптомов рака желудка не существует. Выраженные симптомы появляются, когда опухоль...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Величина мембранного потенциала является отрицательной, т.е. внутреннее содержимое – отрицательно относительно внешней среды и зависит от скорости диффузии ионов. Основным потенциалобразующим ионом является ион калия, вернее, его поток из клетки наружу. Ионы натрия и хлора могут его изменять. Так, мембранный потенциал кардиомиоцитов рабочего миокарда, почти чисто калиевый и его величина равна –90мВ; у скелетной мышцы его величина составляет также -90мВ, но в его формировании принимают участие входящий медленный ток ионов натрия (деполяризующий), входящий ток ионов хлора (гиперполяризующий) и более сильный выходящий калиевый ионный ток. Мембранный потенциал покоя аксона равен –70мВ из-за подключения к выходящему калиевому току медленного и небольшого входящего тока ионов натрия.
2.2.4.1.Изменения мембранного потенциала. Изменения мембранного потенциала, вызываемые электрическим током, подразделяют на пассивные и активные.
Пассивные, или электротонические изменения определяются пассивными электрическими характеристиками как самой мембраны, так и всей клетки (волокна) в целом. Такими характеристиками являются омическое сопротивление и емкостное сопротивление клетки или волокна, чисто физические характеристики мембраны как проводника для электрического тока.
Электротонические изменения были открыты в 1859 году Пфлюгером. Он установил, что при пропускании постоянного тока в момент замыкания электрической цепи пороги раздражения в области катода понижаются, а в области анода возрастают. Эти изменения Пфлюгер назвал катэлектротоном и анэлектротоном, в целом они получили названия физиологического электротона. Исследования Б.Ф.Вериго (1883,1888,1901) существенно дополнили данные, полученные Пфлюгером. Было показано, что начальное катэлектротоническое повышение возбудимости при продолжающемся действии тока постепенно сменяется понижением возбудимости. Это понижение возбудимости получило название «катодическая депрессия». В области анэлектротона снижение возбудимости при размыкании цепи электрического тока становится менее выраженным и переходит в анодическую экзальтацию.
|
|
Пассивные сдвиги мембранного потенциала возникают при действии на возбудимые образования электрического тока любой силы, формы или направления. Однако, если при действии гиперполяризующего (анодного) и слабого деполяризующего (катодного) тока пассивные изменения потенциала могут наблюдаться в чистом виде, то при близких к порогу и сверхпороговых деполяризующих стимулах они сопровождаются активными сдвигами потенциала; локальным ответом и потенциалом действия, которые обязательно связаны с изменениями ионной проницаемости мембраны (рис.6).
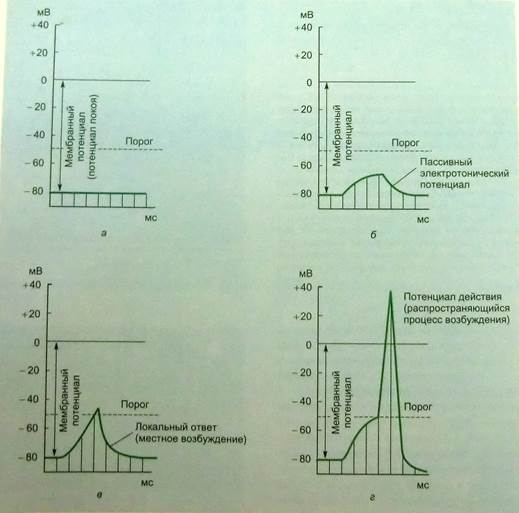
Рис.6 Изменение мембранного потенциала в зависимости от силы раздражения. (Рис. взят из кн.: Фунд. и клин. физиол.М.,2004. Под ред. А.Камкина и А.Каменского)
Локальный ответ. Пассивные, электротонические изменения мембранного потенциала, вызываемые деполяризующим током, при приближении его силы к порогу порождают активную подпороговую электрическую реакцию – так называемый локальный ответ.
Свойства локального ответа. Свойства локального ответа изучались в двух планах. С одной стороны, выявляли его отличия от электротонического потенциала, с другой стороны – сопоставляли локальный ответ и потенциал действия. По своим свойствам локальный ответ очень существенно отличается от пассивного электротонического потенциала:1) в то время как амплитуда электротонического потенциала прямо пропорциональна силе тока, локальный ответ находится в нелинейной зависимости от этой силы; 2)при раздражении однородного участка мембраны электротонический потенциал нарастает и спадает по экспоненциальной кривой со скоростью, которая при данной силе тока определяется физическими свойствами мембраны (R – омическое сопротивление, C – емкость мембраны; RC – постоянная времени). Локальный ответ возрастает по S- образной кривой.
|
|
Сходство локального ответа и электротонического потенциала ограничивается лишь тем, что как тот, так и другой, т находятся в градуальной зависимости от силы стимула (но в одном случае она линейна, в другом – не линейна). Во время развития катэлектротонического потенциала, возбудимость нервного волокна повышается.
Локальный ответ по ряду свойств сходен с потенциалом действия.
1). Он способен к самостоятельному развитию: сначала к нарастанию, а затем к снижению после окончания вызвавшего его короткого стимула.
2). Во время локального ответа, так же как и при генерации потенциала действия, происходит снижение общего сопротивления мембраны (импеданса), т.е., повышение ионной проницаемости.
3). В последействии локального ответа обнаруживается фаза снижения возбудимости, которая по своей динамике очень сходна с периодом относительной рефрактерности, сопровождающим потенциал действия.
Вместе с тем, локальный ответ а) не имеет четкого порога возникновения; б) не сопровождается абсолютной рефрактерностью, напротив, возбудимость во время локального ответа обычно повышается; в) локальный ответ способен к суммации при применении второго подпорогового стимула на фоне возникновения локального ответа от предыдущего раздражения; г) не подчиняется правилу «все или ничего» - его амплитуда и крутизна нарастания градуально увеличивается как при повышении силы подпорогового тока, так и при повторном ритмическом раздражении нервного волокна подпороговыми стимулами редкой частоты. Однако, эти четкие грани, существующие между потенциалом действия и локальным ответом, частично или полностью стираются при различных воздействиях, угнетающих процесс регенеративной деполяризации мембраны. При небольшом катэлектротоне в начальную стадию воздействия на волокно избытка ионов калия, новокаина, уретана, анизотонических растворов, эзерина, простигмина нервное волокно утрачивает способность к генерации нормальных потенциалов действия. Вместо них возникают ответы, которые по своим свойствам занимают промежуточное положение между потенциалом действия и локальным ответом.
|
|
2.2.5.Потенциал действия. К истории вопроса. Современные представления о природе потенциала действия. Основоположник современной мембранной теории Бернштейн (1902,1912) рассматривал потенциал действия как «отрицательное колебание» потенциала покоя. Он полагал, что при раздражении нервного или мышечного волокна в точке приложенного стимула происходят какие-то химические изменения, вызывающие обратимое повышение ионной проницаемости клеточной мембраны. Мембрана утрачивает свою избирательную проницаемость для ионов калия, вследствие чего разность потенциалов, имевшая место в покое, быстро выравнивается, и потенциал покоя падает до нуля. Последующее восстановление исходной ионной проницаемости приводит к возвращению мембранного потенциала к уровню потенциала покоя. После исследований Бернштейна было сделано много попыток экспериментального изучения изменения ионной проницаемости при возбуждении. Люллиес (1936), Кол, Куртис (1936) исследовали продольный импеданс (общее сопротивление) седалищного нерва лягушки и обнаружили его снижение во время проведения нервных импульсов. Аналогичные изменения были получены Куртисом, Колом (1938) и для поперечного импеданса гигантского аксона кальмара. Исследования Тасаки (1953) на одиночном перехвате Ранвье нервных волокон лягушки дали прямое доказательство предположению Бернштейна о повышении ионной проницаемости мембраны при возбуждении. Оставалось, однако, неясным для каких именно ионов возрастает эта проницаемость. Бернштейн полагал, что при возбуждении происходит увеличение проницаемости мембраны для всех ионов. Поэтому потенциал действия он рассматривал лишь как кратковременное исчезновения потенциала покоя, тем более неожиданным оказалось открытие факта превышение потенциала действия над потенциалом покоя. Данные измерения были сделаны вначале с помощью внеклеточных отведений в опытах на гигантском аксоне кальмара (Ходжкин, Хаксли.1939), а затем с применением внутриклеточных электродов (Ходжкин, Хаксли,1939;Кол, Куртис,1939). Разработка метода измерения мембранного потенциала с помощью стеклянных внутриклеточных микроэлектродов (Грэхем, Герард, 1946; Линг, Герард,1949) позволила выявить превышение потенциала покоя практически у всех возбудимых клеток. Миелинизированные нервные волокна быстро повреждаются при введении в них микроэлектрода, поэтому для измерения абсолютных величин потенциалов покоя в этих возбудимых образованиях была разработана специальная методика, позволяющая отводить эти потенциалы от одиночного перехвата Ранвье с помощью обычных внеклеточных электродов (Хаксли,Штемпфли, 1951; Франкенхойзер, 1957). Было установлено, что потенциал покоя перехвата Ранвье хорошо препарированного нервного волокна составляет, примерно,
|
|
-70÷ -75мВ, а амплитуда потенциала действия достигает +115÷ +120 мВ. Дальнейшие исследования с применением перфузии гигантского аксона кальмара, волокон скелетной мышцы и других возбудимых образований с различной концентрацией ионов натрия, ионов калия и других ионов показали зависимость крутизны восходящей фазы потенциала действия от концентрации ионов натрия в наружном растворе. Приведенные опыты подтвердили предположение о том, что возникновение потенциала действия связано с повышением натриевой проницаемости. Теоретические расчеты показали, что для получения потенциала действия амплитудой 100мВ необходимо, чтобы натриевая проницаемость мембраны возросла в 500 раз: от 0,04 до 20. Полученные данные поставили вопрос о причинах восстановления исходной разности потенциалов при возбуждении и о механизме возникновения следовой гиперполяризации.
Для объяснения этих явлений было сделано предположение, что повышение натриевой проницаемости мембраны является только кратковременным процессом, сменяющимся последующей инактивацией механизма, обеспечивающего перенос ионов натрия через мембрану вдоль электрохимического градиента и, что деполяризация мембраны, наряду с повышением натриевой проницаемости, вызывает постепенно нарастающее увеличение его калиевой проницаемости. Дальнейшее изучение ионных токов при возбуждении с применением методов «фиксации мембранного потенциала», «меченых атомов», а также с использованием блокаторов и активаторов ионной проницаемости доказали роль диффузии ионов натрия в генезе потенциала действия и наличие ионных каналов, по которым осуществляется эта диффузии.
|
|
|

Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций...

Индивидуальные очистные сооружения: К классу индивидуальных очистных сооружений относят сооружения, пропускная способность которых...

Эмиссия газов от очистных сооружений канализации: В последние годы внимание мирового сообщества сосредоточено на экологических проблемах...

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!