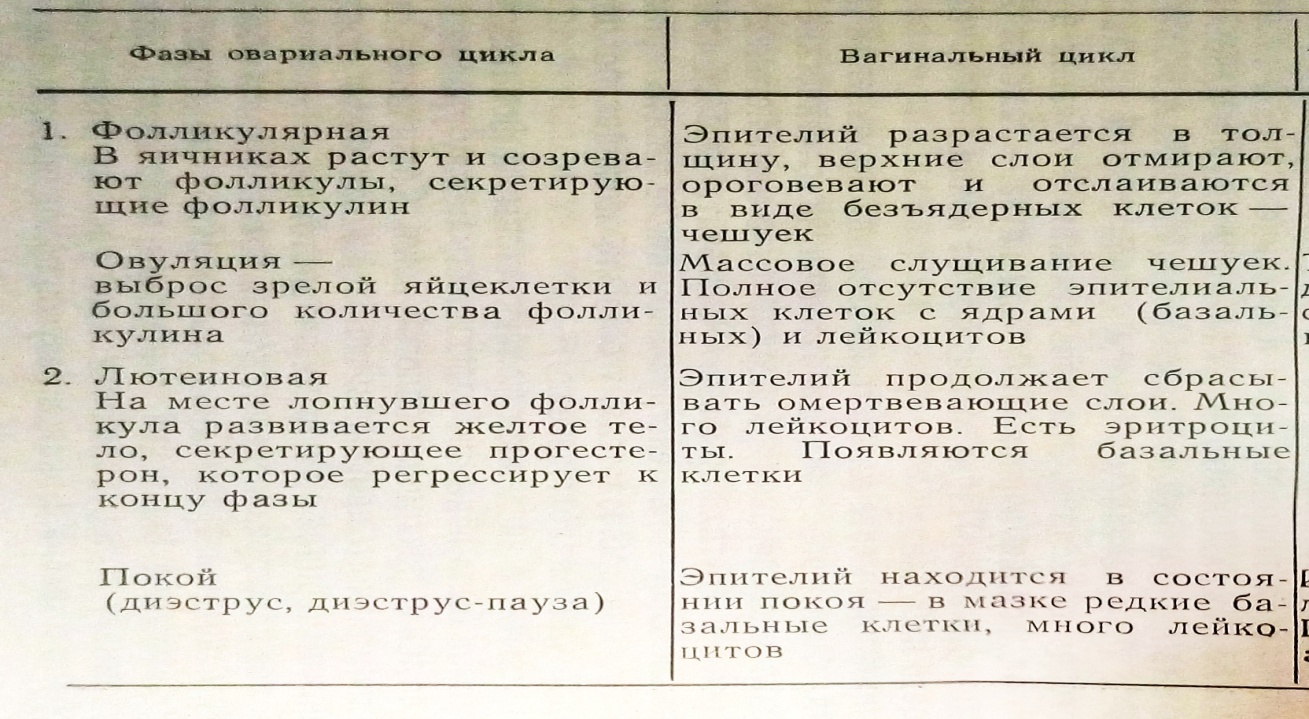
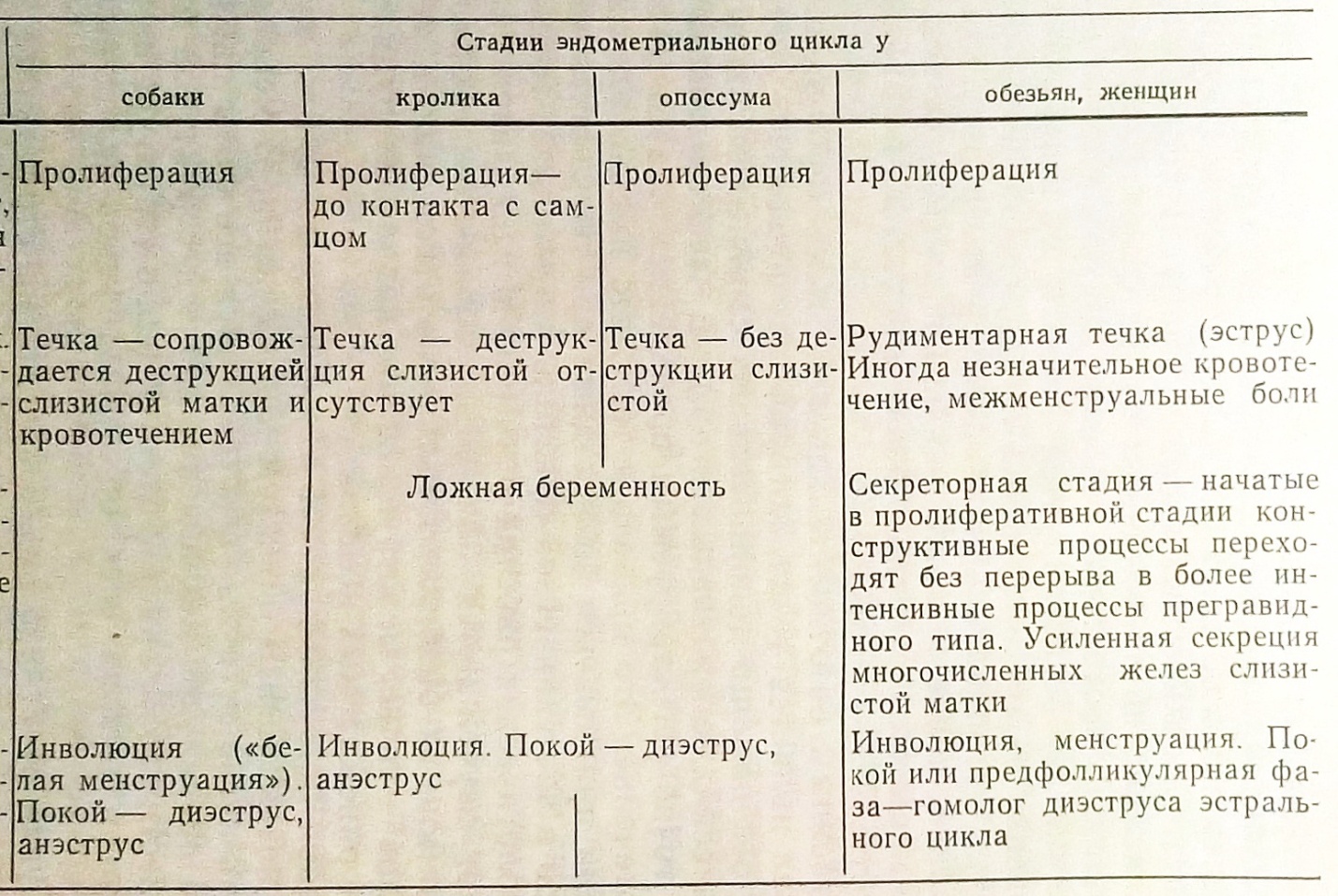
Общими для циклов всех млекопитающих животных оказываются процессы, происходящие в яичниках и составляющие овариальный цикл. Сходно развиваются и конструктивные процессы эндометрия. Различия сводятся к неодинаковым по интенсивности и продолжительности морфофизиологическим изменениям у разных представителей млекопитающих животных.
Нет принципиальных различий и в самой общей характеристике секреторной стадии эндометрия, которая заключается в подготовке к возможной беременности, и в инволюции развившихся изменений в случаях, если оплодотворения не произошло. При ложной беременности «простых» и «двойных» циклов, когда желтое тело менее активно, чем при истинной беременности, секреторные изменения эндометрия не достигают большой интенсивности. В менструальных циклах, безотносительно к беременности, усиленные конструктивные процессы приводят в секреторной стадии к разделению мукозы матки на функциональный и базальный слои, с чем связано и менструальное кровотечение, представляющее собою специфическую надстройку менструальных циклов.
Большим количеством экспериментальных наблюдений, проведенных над течением половых циклов самок приматов, окончательно установлено, что менструальное кровотечение - последняя стадия менструального цикла, завершающая возвращение половой системы к состоянию покоя.
Овуляция происходит обычно (если не считаться с возможностью дополнительных овуляций) в конце пролиферативной стадии. У животных до-приматов она сопровождается течкой, от которой в менструальном цикле приматов остался лишь незначительный рудиментарный след в виде межменструальных кровотечений. У макаков резусов они отмечаются, по К. Гартману, в 90 циклах из 100, у павианов и шимпанзе реже, у человека очень редко и выражаются синдромом межменструальных болей. У остальных животных течка в большинстве циклов сопровождается значительной деструкцией эндометрия матки. Только в «простых» циклах сумчатых и мышевидных грызунов (циклы изолированных самок) овуляция и течка не сопровождаются деструкцией маточного эндометрия.
Гомологом менструального кровотечения у животных можно считать деструктивные изменения эндометрия в конце секреторной фазы, которые достигают значительного выражения и сопровождаются обильным выделением слизи и чешуек вагинального эпителия. У собак, например, выделения такого рода настолько обильны, что их называют «белой менструацией».
19
В половых циклах приматов отмечается еще один тип эндометриальных кровотечений - знак плаценты, соответствующий периоду имплантации яйцеклетки, т. е. 3-4-ой неделе после начала прегравидного цикла; таких кровотечений нет у остальных млекопитающих.
Следует остановиться еще на одной стадии половых циклов, претерпевшей значительные изменения в ходе становления структуры менструальных циклов. Это стадия покоя, или диэструс, астральных циклов животных. У животных эта стадия может быть непродолжительной, если речь идет о циклах, следующих друг за другом, например, в течение брачного сезона, или длительной (анэструс), соответствующей покою половой функции между брачными периодами. В менструальных циклах обезьян, следующих один за другим на протяжении всего детородного периода жизни самки, стадия покоя, или диэструс-пауза, обычно располагается перед стадией пролиферации, в яичниках ей соответствует предфолликулярная фаза. Эта фаза в цикле у обезьян отличается значительными колебаниями продолжительности (от минусовых значений, когда очередная пролиферация начинается в дни менструации, до недель и даже месяцев, переходя в этих условиях в аменорею). В структуре половых циклов у женщин предфолликулярная фаза также имеется, и продолжительность ее весьма изменчива. Наличие этой стадии можно определить с помощью цитологических и гормональных методов исследования. Установление этой стадии в структуре женского менструального цикла помогло понять причины колебаний продолжительности половых циклов, не связанных с патологией. При некоторых условиях стадия покоя, гомологическая диэструс-паузе эстральных циклов животных, у женщин может значительно удлиняться, и тогда развивается физиологическая аменорея. Ниже мы остановимся на этом явлении подробнее.
Эволюция половых циклов у млекопитающих. Черты сходства в структуре половых циклов млекопитающих животных (включая приматов) не случайны. Они обусловлены генетическим родством, общностью происхождения всего класса. К сожалению, из-за скудности сведений о структуре половых циклов у большинства представителей класса млекопитающих до сих пор еще нельзя представить себе с достаточной полнотой эволюцию половых циклов.
Исходным типом циклов, вероятно, были астральные циклы «простой» формы, которые сохранились до настоящего времени у сумчатых животных. У копытных и хищных животных структура половых циклов усложнилась, и за счет добавления стадии ложной беременности она приобрела «двойную» форму. В отряде грызунов развились как простые циклы (сходные с циклами крольчих), так и полиморфные (циклы мышевидных грызунов: крыса, мышь), которые в зависимости от условий могут иметь «простую» или «двойную» форму с переходом в ложную беременность. Конвергентно почти во всех отрядах животных у одной части сохранилась спонтанная овуляция, у других развилась «зависимая» овуляция (опоссум, кошка, кролик).
Половые циклы приматов развились из эстральных циклов. Они сохранили «простую» форму, спонтанную овуляцию, рудименты течки и диэструса, но в их структуре развились и дополнительные особенности - менструальное кровотечение, знак плаценты.
Изменчивость половых циклов
Мы показали, что структура половых циклов в ряду млекопитающих животных в общем сходна. На этой общей основе развиваются различные типы циклов млекопитающих, характеризующие отдельные группы животных. Структура и тип полового цикла относятся к видовым физиологическим особенностям и могут иметь таксономическое значение. Менее постоянными оказываются степень выраженности внешних проявлений циклов, продолжительность периодов составляющих и всего цикла
20
в целом. Видовые и индивидуальные изменения продолжительности половых циклов до последнего времени не привлекали к себе внимания.
Раньше всего в связи с запросами клиники началось всестороннее изучение половых циклов женщин. Долгое время существовало ложное мнение о постоянстве ритма менструаций. Считалось бесспорным, что циклы у женщин продолжаются 28 дней, так называемый лунный месяц. Возможно, это представление развилось под влиянием «лунной» теории менструаций, которая ставила циклические проявления женского организма в связь с фазами луны.
Лишь в 40-х годах нашего столетия появились первые сведения об аритмичном течении половых циклов у женщин в норме. Сначала аритмичность была признана для молодых женщин, находящихся в периоде становления половой функции. Позже колебания продолжительности половых циклов были доказаны и для взрослых женщин. Хотя у молодых аритмичность, как правило, выражена сильнее. Постепенно, с возрастом, у женщин устанавливается более или менее постоянный ритм. И все же границы фактической изменчивости продолжительности половых циклов могут лежать в пределах от нескольких до десятков и сотен дней. Fluhman (1934) для менструальных циклов здоровых женщин установил пределы от 7 до 256 дней, Bartelmez (1951) -от 11 до 144 дней, Allen (1933) - от 13 до 84 дней и т. п. Как оказалось, лишь незначительная часть циклов имеет продолжительность, равную 20-30 дням (от 9 до 75%, по данным разных авторов).
Распределение женских менструальных циклов по продолжительности совпадает с нормальным. Наиболее частыми оказались варианты в 27-28-29 дней. Обращает на себя внимание асимметрия кривой со сдвигом вправо. Это указывает на то, что имеется больше факторов для удлинения, чем для укорочения половых циклов. Следует отметить, что в пределах границ колебания продолжительности половых циклов, установленных на больших группах женщин, индивидуальные колебания могут иметь свои, более ограниченные и постоянные пределы. Так, отчетливо выделяются группы женщин, имеющих в среднем половые циклы в 21-22 дня, 25-27, 28-30 дней и более продолжительные.
Поскольку единственным бесспорно и легко регистрируемым проявлением цикла у женщин является менструальное кровотечение и прослеживание всей структуры цикла у них недоступно без применения специальных методов обследования, вопрос о том, за счет чего изменяется продолжительность половых циклов у здоровых женщин, долгое время оставался нерешенным.
И так же как вопрос об аритмии менструальных циклов у женщин был поднят и решен после того, как аритмия была установлена на обезьянах, Harlman (1934), так и изменения половых циклов по продолжительности были поняты только после исследований, проведенных на обезьянах.
Мы вернемся к этому вопросу после знакомства с изменчивостью половых циклов у обезьян и других животных.
К сожалению, в литературе почти нет данных о вариабельности половых циклов у самок животных.
Самок белых мышей обследовала А. А. Кудрявцева (см. Алексеева, 1974). Под наблюдением было 10 самок. Вагинальные лаважи брались у них 1-2 раза в день. Для регистрации структуры цикла была применена 4-балльная система (см. рис. 8).
На графиках отчетливо видно изменение продолжительности 100 половых циклов у 10 мышей (по 10 циклов у каждой). Общая продолжительность циклов изменялась от 3 до 20 и более дней. Видно также, что наиболее изменчивой у мышей оказалась стадия диэструса. Сходные данные были опубликованы в 1951 г. И. А. Эскиным. Хотя автор, изображая графически циклы у крыс, выделял только течку, он, отмечая нару-
21
шения половых циклов у крыс под влиянием различных фармакологических воздействий, в тексте указывал, что прежде всего изменяется продолжительность интервала за счет удлинения диэструса. Сходные указании можно «между строк» найти в работах многих авторов экспериментировавших на животных. Но мы не встретили работ, где бы принималась во внимание продолжительность составляющих цикл стадий, за исключением течки, время наступления и продолжительность которой обычно фиксируется.
Половые циклы и их изменчивость у самок обезьян. Половая функция обезьян изучалась многими авторами, и можно без преувеличения сказать, что половые циклы обезьян могут служить естественной моделью половых циклов женщин. Сходство структуры, выраженности различных проявлений течения отдельных фаз цикла, изменения по продолжительности отдельных фаз и всего цикла в целом все это дало основания, с одной стороны, использовать половые циклы обезьян в качестве экспериментальной модели менструального цикла женщин, с другой стороны, сделало правомочным перенос данных, полученных в процессе изучения половой функции обезьян, на человека.
Половые циклы макаков резусов впервые стал изучать Hartman (1932). Он обследовал 712 циклов у 42 самок колонии Карнеги и отметил большую вариабельность продолжительности циклов - от 9 до 200 дней, что и послужило основанием выступить против общепринятого в те годы представления о постоянстве продолжительности половых циклов у женщин. Но Hartman не проанализировал причин изменения продолжительности половых циклов у макаков. В поисках нормы он просто отбросил все значения циклов, превышавшие 50 дней, и, оставив 392 цикла от 22 наиболее «благополучных» самок колонии, получил среднюю продолжительность циклов в 32,5 дня при Моде=28 дней. Это привело к удивительному сообщению Hartman о сходстве по продолжительности половых циклов самок макаков с менструальными циклами женщин, в то время как другими авторами единодушно отмечалась большая продолжительность половых циклов у всех других видов обезьян, даже таких близких человеку, как шимпанзе.
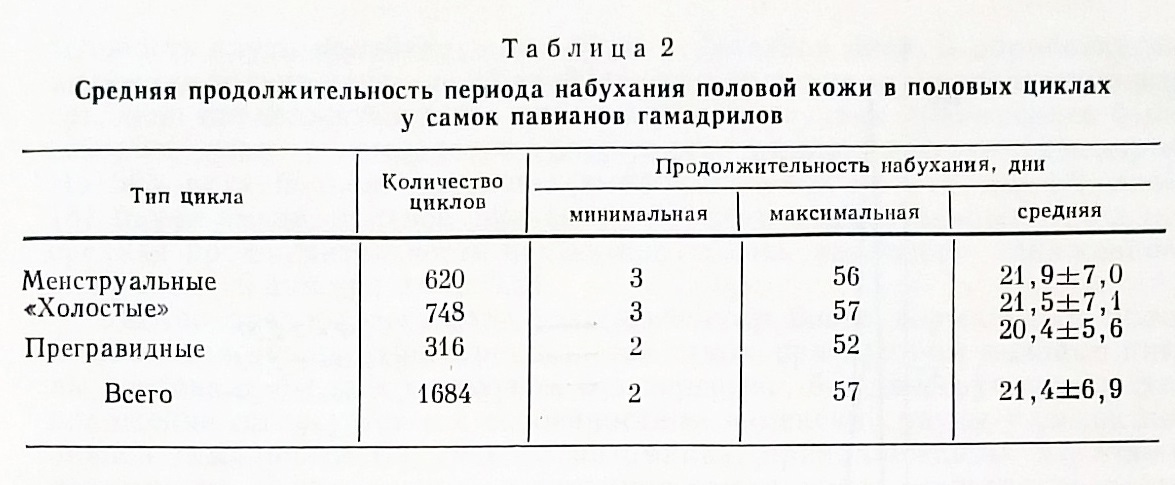
Половые циклы и их изменчивость у самок павианов гамадрилов. В условиях Сухумского питомника нами было проведено изучение 1684 половых циклов у 87 самок павианов гамадрилов. Как оказалось, непременным внешним выражением цикла у павианов было набухание половой кожи. Остальные части цикла могли развиваться по-разному, в зависимости от судьбы фолликула в данном цикле. Если имела место овуляция, то после отбухания половой кожи сразу или через несколько дней наступала менструация. После нее в течение нескольких дней половая кожа оставалась в покое, а затем развивалось новое набухание. В таком полноценном цикле регистрировались все четыре фазы. Если цикл был ановуляторным, «холостым», то после отбухания половая кожа оставалась в покое, а затем снова начиналось набухание следующего цикла. Часть циклов у павианов переходила в беременность, и в этом случае можно было наблюдать только набухание - отбухание половой кожи и далее - длительный период ее покоя на время беременности (около 6 месяцев). Из общего числа обследованных циклов 617 были менструальными, 748 - «холостыми», т.е. безменструальными, 316 - закончились беременностями.
Анализируя продолжительность набухания-отбухания половой кожи во всех циклах, мы обнаружили, что продолжительность набухания оказалась одинаковой в среднем в циклах всех типов (табл 2). Очевидно, следует предположить, что продолжительность набухания, а вместе с ним и пролиферативной фазы цикла не зависит у павианов от судьбы цикла, от характера его второй части - будет ли она прегравидной или секреторной.
22
Установлена линейная зависимость между продолжительностью набухания половой колеи (b) и днем максимального набухания (а), а именно а = 0,635 b. Или: пролиферативная фаза цикла пропорциональна длительности набухания в днях и составляет 0,635 последнего (Бочкарев, 1935).
Поскольку средняя продолжительность периода набухания-отбухания половой колеи для всей совокупности цикла была определена в 21.4 дня, средняя теоретическая продолжительность пролиферативной фазы цикла павианов гамадрилов составила 0,635х21,4= 13,6 дня.
Вторая регистрируемая часть полового цикла - покой половой колеи перед менструацией, или интерменструм, была прослежена в 620 циклах у 87 самок и оказалась равной в среднем 8,4 дня при ס = ±5,42 с колебаниями в пределах от 3 до 30 дней. Зная эту величину, можно вычислить величину секреторной фазы в цикле павианов. Она составляется из части отбухания половой колеи после дня овуляции и предменструального периода покоя половой колеи. Весь период набухания продолжался в среднем 21,4 дня. Из них на долю пролиферативной фазы пришлось
13.4 дня, а для секреторной осталось 21,4-13,4=8,0 дня. Следовательно, общая продолжительность секреторной фазы должна равняться 8,0+8,4=16,4 дня. Как и в цикле женщин, секреторная фаза оказалась более длительной, чем пролиферативная.
Продолжительность менструального кровотечения исчислялась в целых днях. Этот период цикла был наименее изменчив. Для 620 циклов он в среднем был равен 2,5 дня при ס = ±0,45 (пределы колебаний 1-7 дней).
Среднюю теоретическую продолжительность половых циклов павианов гамадрилов Сухумского питомника получили, сложив вычисленные средние значения всех фаз цикла: 13,4+16,4 + 2,5=32,3 дня.
Вычисленная для 613 циклов фактическая средняя продолжительность оказалась несколько большей: М = 33,1 дня при ס = ±9,02. При этом за пределами М±3 סосталось 14 циклов. Один из них был 6-дневным, остальные - более 66 дней, т. е. превышали удвоенное среднее значение.
Между пролиферативной и секреторной фазами цикла был вычислен коэффициент корреляции, он оказался меньше 1. Можно с большим основанием говорить, что нормальная изменчивость продолжительности менструального цикла, по-видимому, обусловлена комбинированием независимых колебаний продолжительности фаз цикла, вариабельность которых в норме невелика (в пределах М±3 ס).
Однако если продолжительность половых циклов у саамок павианов отсчитывать без учета структуры цикла, а просто от одной менструации до другой, то общая продолжительность циклов получится значительно большей, и эта разница будет образовываться за счет вариабельности предфолликулярной фазы, или паузы. На нашем материале продолжи-
23
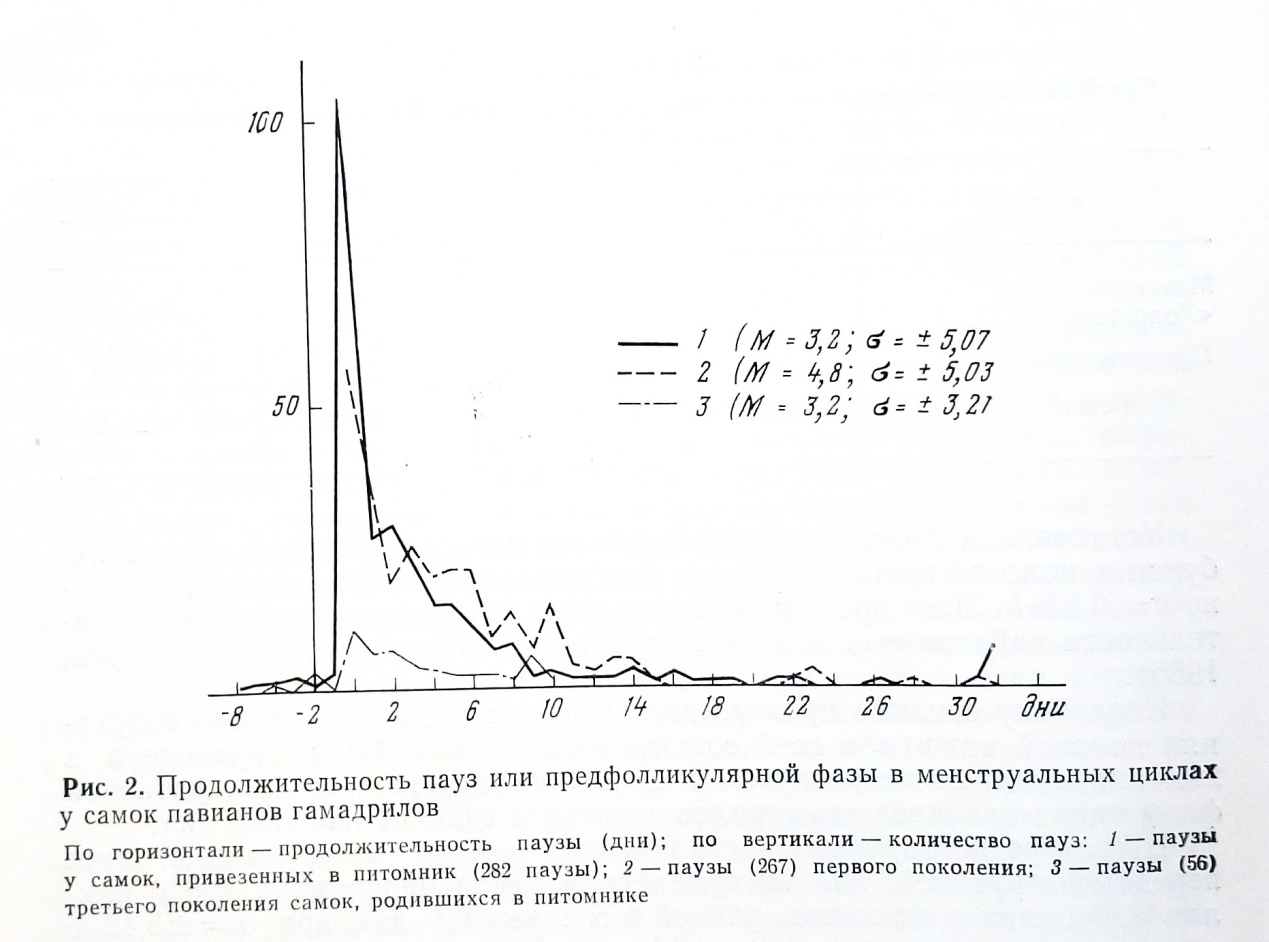
тельность паузы колебалась от -8 до + десятков дней. В обработку мы включили только паузы до 33 дней, так как большие значения превышали среднюю продолжительность цикла и в этих случаях правомернее было говорить уже о выпадении очередного цикла, или об аменорее. Из 605 пауз больше половины имели значение от -8 до +2 дней, 181 пауза имела нулевое значение. За счет отрицательных значений средняя продолжительность паузы получилась несколько заниженной она равна 3,9 дня при ס = ±5,24.
Можно предполагать, что физиологически более нормальным положением для циклов павианов является такое, при котором половые циклы следовали бы друг за другом без перерыва, без диэструс-паузы. Это положение согласуется и с особенностями изменений паузы у самок павианов гамадрилов Сухумского питомника, принадлежащих к разным поколениям. У привезенных в питомник самок паузы встречались почти во всех циклах и были более продолжительными. У самок, рожденных в питомнике, величина пауз в первом поколении увеличилась, а в следующем снова начала уменьшаться (рис. 2).
На основании всего изложенного мы считаем, что следует различать истинную вариабельность продолжительности менструальных циклов, когда удлинение идет за счет изменений активных периодов цикла - пролиферации, секреции и менструации, и мнимую вариабельность, обусловленную изменениями продолжительности неактивной части цикла - предфолликулярной фазы, или паузы. Мнимая вариабельность намного превышает истинную и маскирует ее.
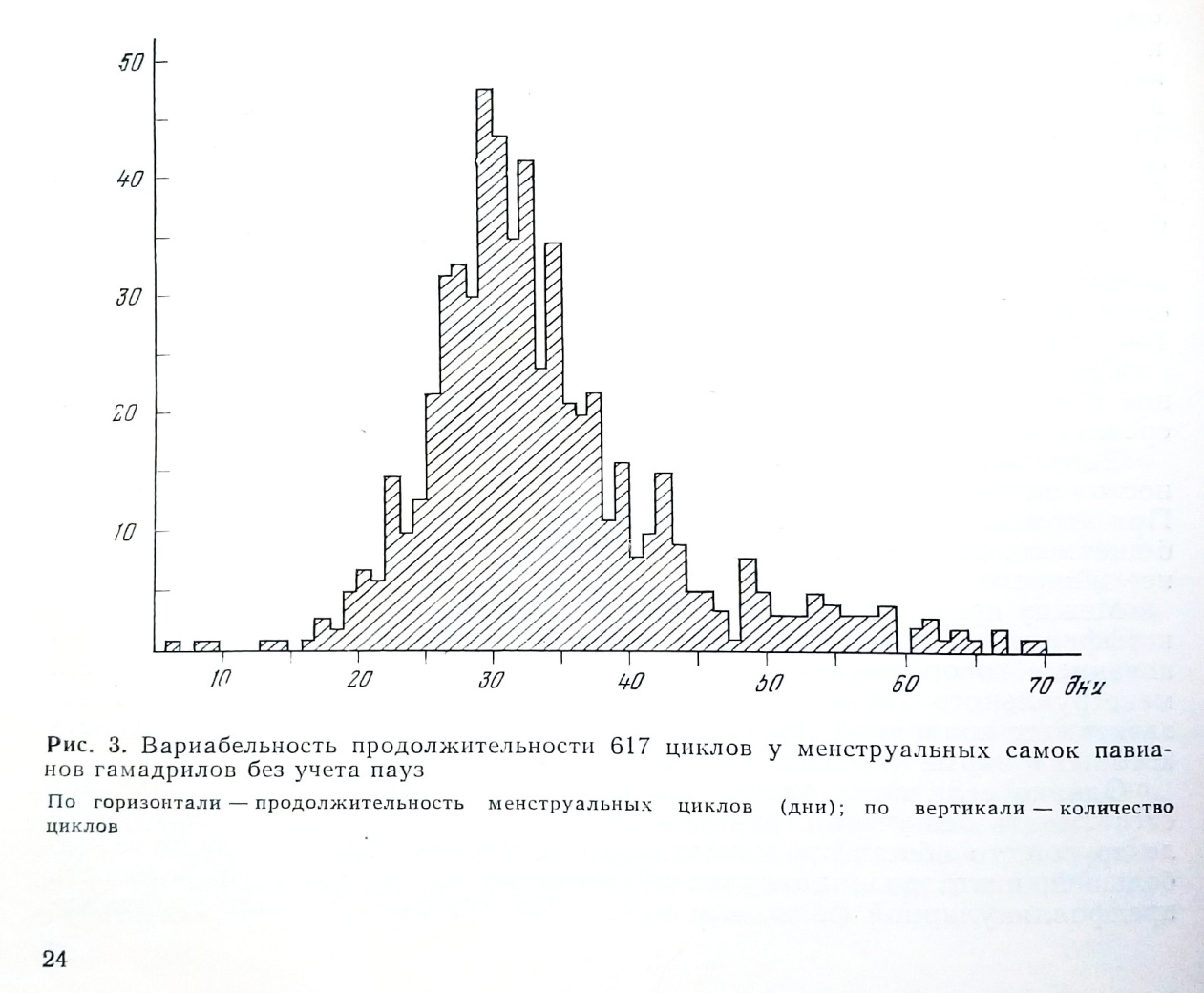
Поэтому истинную продолжительность цикла можно получить, отбросив значения паузы (рис. 3). На гистограмме продолжительности половых циклов павианов гамадрилов Мода = 29 дням. Правая ветвь более полога, что свидетельствует о тенденции циклов к удлинению. Если включить значения паузы, то Мода передвинется на значения 35-36 дней, а тенденция к удлинению циклов будет выражена еще ярче за счет специфических особенностей паузы.
Сопоставляя эти данные с полученными Hartman, мы видим, что вычисленная без учета паузы средняя продолжительность половых циклов павианов также сближается со средней продолжительностью половых циклов макаков и человека. Тем самым установленное Hartman противопоставление продолжительности половых циклов макаков всем остальным видам приматов оказывается несостоятельным по отношению к павианам, в частности.
Обширные данные имеются в литературе о половых циклах шимпанзе. Yong, Yerkes (1943) описали их циклы, отметив большие и хорошо развитые набухания половой кожи, которые позволяли точно регистр ц- ровать структуру циклов. Авторы выделили фазу покоя половой кожи перед набуханием и назвали ее «preswelling phase». Они отметили продолжительность этой фазы наряду с другими фазами цикла в каждом цикле у каждой самки и включили ее в продолжительность цикла. При этом авторы получили среднюю продолжительность циклов на неделю больше, чем у макаков и павианов. Если же пересчитать данные Yong и Yerkes, не принимая во внимание значение пауз, то и у самок шимпанзе средняя продолжительность циклов уменьшится до 28,5 дня при Моде=29 дням. Из работы этих авторов с очевидностью следует, что, как правило, у шимпанзе удлинение циклов происходит за счет паузы, или предфолликулярной фазы. Этот вывод подтверждается и в работе Erikson (1963). Автор тщательно изучал половые циклы у самки шимпанзе. Он пользовался методом вагинальных мазков, регистрацией всех внешних проявлений цикла и оценкой величины набухания по объемным муляжам. Два последовательных цикла продолжались 41-46 дней. При этом секреторная фаза составляла 12 и 15 дней, а пролиферативная (вместе с паузой) 26 и 34 дня. В третьем цикле самка
25
была овариектомирована, когда набухание достигло максимума. В яичниках у шимпанзе впервые было обнаружено свежее желтое тело, которое автор и описал, отметив большое сходство с желтым телом яичника женщин. Половые циклы у двух самок гиббонов колонии на о. Сант-Яго наблюдал и описал Carpenter (1941). За 1-й день цикла он принимал начало менструального кровотечения. Для 17 половых циклов гиббонов средняя продолжительность оказалась равной 29,7 дня при ס = ±4,l дня и границах колебаний от 21 до 43 дней.
К сожалению, в литературе имеется мало сведений о половых циклах других видов обезьян. Но сходство структуры цикла и общность причин удлинения циклов в норме, установленная для всех изученных приматов (макаков и павианов нескольких видов, шимпанзе, гиббонов и человека), позволяет считать эти закономерности общими для всех видов приматов. Видовые различия сводятся к вариациям степени внешних проявлений цикла (наличие или отсутствие половой кожи, течки, менструации).
Открытие в структуре цикла приматов предфолликулярной фазы (гомолога диэструса у современных грызунов) помогло понять некоторые особенности вариабельности женских половых циклов по продолжительности.
Широкую известность приобрели работы Ogino (1930) и Knauss (1934), которые путем большого количества лапаратомий установили, что овуляция у женщин в большинстве случаев наступает за 12-16, чаще за 14 дней до начала менструации. Из обследованных Огино пациентов только у 20% отмечались регулярные 23-29-дневные циклы. У остальных 80% продолжительность циклов колебалась как в сторону 21-22 дней, так и удлинялась до 40-45 и более дней. Поэтому оказалось особенно интересным постоянство дня овуляции по отношению к окончанию цикла, проявившееся, несмотря на нерегулярность последних. Согласно вычислениям Огино - Кнаусса, общая продолжительность полового цикла женщин зависит от вариабельности по продолжительности его первой половины, тогда как вторая, соответствующая секреторной стадии в норме, почти не изменяется.
Представляется маловероятным, чтобы у здоровой женщины фолликулярная фаза, во время которой начинается рост яйцеклетки и происходит созревание фолликула, могла значительно изменять свою продолжительность - от 9 дней в 21-дневном цикле до 20-25 и более дней при циклах в 35-40 дней и далее. Если бы структуру цикла у женщин можно было бы так же легко прослеживать, как в циклах обезьян, обладающих половой кожей, очевидно, предфолликулярная фаза давно была бы выделена и стало бы ясным, что именно ее изменения определяют во многих случаях аритмичность, свойственную половым циклам здоровых женщин. Вероятно, по аналогии с циклами других приматов пауза у женщин также стремится при нормальной жизнедеятельности к нулевому значению. Однако пауза легко приобретает положительную продолжительность и, удлиняясь, изменяет общую продолжительность цикла. Теоретически можно рассчитать длительность отдельных стадий женского полового цикла. При 28 29-дневном цикле 14 дней приходятся на секреторную фазу (по Огино-Кнауссу). Остальные 14-15 дней составляются из 3 4 дней менструации, 7 8 дней пролиферативной стадии и 3-4 дней паузы.
О том, что в структуре половых циклов у женщин действительно имеется период паузы, или предфолликулярная фаза свидетельствуют наблюдения П. В. Бочкарева (1937), который у многих женщин отмечал пониженную десквамацию вагинального эпителия растягивающуюся после менструации при удлиненных циклах на много дней пока не начинался нормальный цикл. Сходные данные приводил в своей работе и И. Д. Арист (1940). Многочисленные литературные данные были суммированы А. А. Машковцевым, которые говорят о том что у женщин в
26
яичниках после окончания цикла (после менструации) фолликулы обычно не превышают определенного диаметра и могут находиться в таком состоянии неопределенно долгое время, а затем начинают расти, что является началом нового цикла.
Убедительные подтверждения наличия стадии относительного покоя яичников после окончания одного и до начала другого цикла были получены путем прямых определений количества половых гормонов в моче у женщин. Начиная с С. Е. Файермарк (1945), определявшей содержание фолликулина в моче биологическим путем, и в многочисленных работах последующих лет, в которых приводятся результаты определения половых гормонов на протяжении всех фаз циклов, почти в каждом случае после окончания менструации экскреция эстрогенов некоторое время колеблется, не превышая некоего низкого уровня, и лишь позже начинает увеличиваться. Чем более длителен цикл, тем продолжительнее этот период низкого уровня экскреции эстрогенов, т. е. продолжительнее пауза (предфолликулярная фаза).
При определении эстрогенов в цикле у павианов мы в период низкого уровня экскреции не отмечали активности половой кожи. Набухание начиналось только тогда, когда уровень экскреции поднимался выше порогового.
В заключение следует отметить, что половые циклы грызунов, обезьян и человека весьма изменчивы по продолжительности даже в нормальных условиях существования и при отсутствии патологии. Однако эта изменчивость преимущественно создается за счет изменения продолжительности не активной части цикла, а периодов диэструса и его гомолога в половых циклах приматов - предфолликулярной фазы, или паузы. Собственно цикл, его активная часть, слагающаяся из процессов роста и развития фолликулов и желтых тел, процессов достаточно детерминированных и определенных, также может изменяться во времени, но в значительно более узких пределах.






