Углеводы
В мозговой ткани имеются гликоген и глюкоза. Общее содержание глюкозы в головном мозге разных животных составляет в среднем 1–4 мкмоль на 1 г ткани, а гликогена – 2,5–4,5 мкмоль на 1 г ткани.
В мозговой ткани имеются также промежуточные продукты обмена углеводов: гексозо- и триозофосфаты, молочная, пировиноградная и другие кислоты.
Таблица 19.3. Средние данные о содержании некоторых метаболитов обмена углеводов в головном мозге крыс
| Метаболит
| Содержание,
мкмоль на 1 г
сырой массы ткани
| Метаболит
| Содержание,
мкмоль на 1 г
сырой массы ткани
|
| Глюкозо-6-фосфат
Фруктозо-6-фосфат
Фруктозо-1,6-бисфосфат
Диоксиацетонфосфат
Глицеральдегид-3фосфат
| 0,039-0,049
0,017-0,023
0,010-0,017 0,024
0,021-0,046
| 3-Фосфоглицерат 2-Фосфоглицерат
Фосфоенолпируват Пируват
Лактат
| 0,085-0,100
0,010-0,016
0,035-0,097
0,120-0,190
1,26-1,70
|
ОСОБЕННОСТИ МЕТАБОЛИЗМА НЕРВНОЙ ТКАНИ Дыхание
Потребление кислорода головным мозгом в состоянии физического покоя достигает 20–25% от общего потребления его всем организмом, а у детей в возрасте до 4 лет мозг потребляет даже 50% кислорода, утилизируемого всем организмом.
Газообмен мозга значительно выше, чем газообмен других тканей, в частности он превышает газообмен мышечной ткани почти в 20 раз. Интенсивность дыхания для различных областей головного мозга неодинакова. Особенно интенсивно расходуют кислород клетки коры мозга и мозжечка.
Поглощение кислорода головным мозгом значительно меньше при наркозе,интенсивность дыхания мозга возрастает при увеличении функциональной активности.
Метаболизм углеводов
Основным субстратом дыхания мозговой ткани является глюкоза.
В физиологических условиях роль пентозофосфатного пути окисления глюкозы в мозговой ткани невелика, этот путь окисления глюкозы присущ всем клеткам головного мозга. Образующаяся в процессе пентозофосфатного цикла восстановленная форма НАДФ (НАДФН) используется для синтеза жирных кислот и стероидов.
1 мин тканью мозга окисляется 75 мг глюкозы.
Основным субстратом дыхания головного мозга является глюкоза крови.
При недостаточном поступлении глюкозы из крови гликоген головного мозга является источником глюкозы, а глюкоза при ее избытке – исходным материалом для синтеза гликогена. Распад гликогена в мозговой ткани происходит путем фосфоролиза с участием системы цАМФ.
Наряду с аэробным метаболизмом углеводов мозговая ткань способна к довольно интенсивному анаэробному гликолизу.
Метаболизм аминокислот и белков
Общее содержание аминокислот в ткани мозга человека в 8 раз превышает концентрацию их в крови. Аминокислотный состав мозга отличается определенной специфичностью. В мозге содержится ряд свободных аминокислот, которые лишь в незначительных количествах обнаруживаются в других тканях млекопитающих. Это γ-аминомасляная кислота, N-ацетиласпарагиновая кислота и цистатионин.
Прежде всего пул свободных аминокислот используется как источник «сырья» для синтеза белков и биологически активных аминов. Одна из функций дикарбоновых аминокислот в головном мозге – связывание аммиака, освобождающегося при возбуждении нервных клеток.
Однако в разных отделах головного мозга скорость синтеза и распада белковых молекул неодинакова. Белки серого вещества полушарий большого мозга и белки мозжечка отличаются особенно большой скоростью обновления. В участках головного мозга, богатых проводниковыми структурами – аксонами, скорость синтеза и распада белковых молекул меньше.
При действии на организм животных возбуждающих агентов в головном мозге усиливается интенсивность обмена белков. Под влиянием наркоза скорость распада и синтеза белков снижается.
Возбуждение нервной системы сопровождается повышением содержания аммиака в нервной ткани.
Метаболизм липидов
Липиды составляют около половины сухой массы головного мозга. Как отмечалось, в нервных клетках серого вещества особенно много фосфоглицеридов, а в миелиновых оболочках нервных стволов – сфингомиелина. Из фосфоглицеридов серого вещества мозга наиболее интенсивно обновляются фосфатидилхолины и особенно фосфатидилинозитол. Обмен липидов миелиновых оболочек протекает с небольшой скоростью. Холестерин, цереброзиды и сфингомиелины обновляются очень медленно.
Ткань головного мозга взрослого человека содержит много холестерина (около 25 г). У новорожденных в головном мозге всего 2 г холестерина; количество его резко возрастает в первый год жизни (примерно в 3 раза), при этом биосинтез холестерина происходит в самой мозговой ткани. У взрослых людей синтез холестерина в головном мозге резко снижается.
Основная часть холестерина в зрелом мозге находится в неэтерифицированном состоянии, эфиры холестерина обнаруживаются в относительно высокой концентрации в участках активной миелинизации. Жирные кислоты образуются в основном из глюкозы, однако частично синтез их происходит из ацетоацетата, цитрата и даже ацетиласпартата.
ЦЕРЕБРОСПИНАЛЬНАЯ ЖИДКОСТЬ
Общий объем цереброспинальной жидкости у взрослого человека в норме составляет около 125 мл; каждые 3–4 ч жидкость обновляется. Состав отличается от состава плазмы крови, что позволяет приписывать сосудистому эндотелию в нервной системе главную роль в осуществлении барьерной функции. Вода в цереброспинальной жидкости составляет 99%, на долю плотного остатка приходится около 1%.
Таблица 19.5. Химический состав цереброспинальной жидкости
| Компоненты
| Содержание
| Компоненты
| Содержание
|
| Белки
Альбумины/глобулины
Остаточный азот
Азот аминокислот
Азот мочевины
Глюкоза
Молочная кислота
Холестерин
| 0,15-0,40 г/л
4:1
8,57-14,28 ммоль/л
1,14-1,93»
2,86-7,14»
2,50-4,16»
1,67 ммоль/л
2,62-5,20 ммоль/л
| Триацилглицерины Лецитин
Na+
К+
Са2+ Сl–
HCO3–
| Следы
»
146 ммоль/л
3,5-4,0»
1,5» 125»
25 »
|
Содержание белка в цереброспинальной жидкости незначительно (0,15–0,40 г/л), причем отношение альбумины/глобулины равно 4; липидов в сотни раз меньше, чем в плазме крови. В ткани мозга, как отмечалось, количество свободных аминокислот велико и во много раз превышает концентрацию их в крови и тем более в цереброспинальной жидкости. Установлено, что некоторые аминокислоты (например, глутаминовая кислота) почти не проникают через гематоэнцефалический барьер. В то же время амиды аминокислот (в частности, глутамин) легко преодолевают этот барьер. Содержание глюкозы в цереброспинальной жидкости относительно велико (2,50–4,16 ммоль/л), но несколько меньше, чем в крови, причем концентрация глюкозы в спинномозговой жидкости может повышаться или снижаться в зависимости от изменений содержания глюкозы в крови.
По содержанию ионов К+ и Na+ цереброспинальная жидкость практически не отличается от плазмы крови. Ионов Са2+ в ней почти в 2 раза меньше, чем в плазме крови. Содержание ионов Сl– заметно выше, а концентрация ионов бикарбоната несколько ниже в цереброспинальной жидкости, чем в плазме.
120. Проведение и передача нервного импульса. Потенциал покоя и потенциал действия. Синапсы, синаптическая передача. Нейротрансмиттеры: ацетилхолин, катехоламины, серотонин, γаминомасляная кислота, глицин, гистамин. Нейропептиды. Механизмы памяти.
Мембрана обладает избирательной проницаемостью: большей для ионов К+ и значительно меньшей для ионов Na+. Кроме того, в нервных клетках существует механизм, который поддерживает внутриклеточное содержание натрия на низком уровне вопреки градиенту концентрации. Этот механизм получил название натриевого насоса.
При определенных условиях резко повышается проницаемость мембраны для ионов Na+.
В состоянии покоя внутренняя сторона клеточной мембраны заряжена электроотрицательно по отношению к наружной поверхности. Объясняется это тем, что количество ионов Na+, выкачиваемых из клетки с помощью натриевого насоса, не вполне точно уравновешивается поступлением в клетку ионов К + . В связи с этим часть катионов натрия удерживается внутренним слоем противоионов (анионов) на наружной поверхности клеточной мембраны. Таким образом, на мембранах, ограничивающих нервные клетки, поддерживается разность электрических потенциалов (трансмембранная разность электрических потенциалов); эти мембраны электрически возбудимы.
При возбуждении, вызванном тем или иным агентом, селективно изменяется проницаемость мембраны нервной клетки (аксона): увеличивается избирательно для ионов Na+ (примерно в 500 раз) и остается без изменения для ионов К + . В результате ионы Na+ устремляются внутрь клетки. Компенсирующий поток ионов К + , направляющийся из клетки, несколько запаздывает. Это приводит к возникновению отрицательного заряда на наружной поверхности клеточной мембраны. Внутренняя поверхность мембраны приобретает положительный заряд; происходит перезарядка клеточной мембраны (в частности, мембраны аксона, т.е. нервного волокна), и возникает потенциал действия, или спайк. Продолжительность спайка не превышает 1 мс. Он имеет восходящую фазу, пик и нисходящую фазу. Нисходящая фаза (падение потенциала) связана с нарастающим преобладанием выхода ионов К+ над поступлением ионов Na+ – мембранный потенциал возвращается к норме. После проведения импульса в клетке восстанавливается состояние покоя. В этот период ионы Na+, вошедшие в нейрон при возбуждении, заменяются на ионы К + . Этот переход происходит против градиента концентрации, так как ионов Na+ во внешней среде, окружающей нейроны, намного больше, чем в клетке после момента ее возбуждения. Переход ионов Na+ против градиента концентрации, как отмечалось, осуществляется с помощью натриевого насоса, для работы которого необходима энергия АТФ. В конце концов все это приводит к восстановлению исходной концентрации катионов калия и натрия внутри клетки (аксона), и нерв готов для получения следующего импульса возбуждения. Заметим, что миелиновые мембраны, образуемые шванновскими клетками, окутывают нервные волокна и служат электрическим изолятором. Этот изоляционный слой покрывает большинство нервных волокон и сильно ускоряет распространение электрической волны (сигнала); при этом ионы входят в клетку и выходят из нее только в тех местах, где изолятор отсутствует. Как уже отмечалось, миелиновая мембрана состоит из фосфолипидов, в частности из сфингомиелина, холестерина, а также белков и гликосфинголипидов.
Потенциал покоя и потенциал действия.
Потенциал покоя- это дефицит положительных зарядов внутри клетки, возникающий за счёт работы натрий-калиевого насоса и (в большей мере) последующей утечки из клетки положительных ионов калия
Потенциал действия- это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
Синапс- узкое пространство (щель), ограниченное с одной стороны пресинаптической, а с другой – постсинаптической мембраной. Пресинаптическая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Постсинаптическая мембрана менее плотная, не имеет отверстий. Подобным образом построены и нервно-мышечные синапсы, но они имеют более сложное строение мембранного комплекса.
В общих чертах картину участия ацетилхолина в осуществлении передачи нервного импульса возбуждения можно представить следующим образом. В синаптических нервных окончаниях имеются пузырьки (везикулы) диаметром 30–80 нм, которые содержат нейромедиаторы. Эти пузырьки покрыты оболочкой, которая образована белком клатрином (мол. масса 180000). В холинергических синапсах каждый пузырек диаметром 80 нм содержит ~ 40000 молекул ацетилхолина. При возбуждении высвобождение медиатора происходит «квантами», т.е. путем полного опорожнения каждого отдельного пузырька. В нормальных условиях под влиянием сильного импульса выделяется примерно 100–200 квантов медиатора – количество, достаточное для инициирования потенциала действия в постсинаптическом нейроне. Деполяризация мембраны синаптических окончаний вызывает быстрый ток ионов Са2+ в клетку. Временное увеличение внутриклеточной концентрации ионов Са2+ стимулирует слияние мембраны синаптических пузырьков с плазматической мембраной и таким образом запускает процесс высвобождения их содержимого. Для выброса содержимого одного пузырька требуется примерно 4 иона Са2+. Выделенный в синаптическую щель ацетилхолин вступает во взаимодействие с белком-хеморецептором, входящим в состав постсинаптической мембраны. В результате изменяется проницаемость мембраны – резко увеличивается ее пропускная способность для ионов Na+. Взаимодействие между рецептором и медиатором запускает ряд реакций, заставляющих постсинаптическую нервную клетку или эффекторную клетку выполнять свою специфическую функцию. После выделения медиатора должна наступить фаза его быстрой инактивации,
или удаления, чтобы подготовить синапс к восприятию нового импульса.
В холинергических синапсах это происходит двумя путями. Первый путь заключается в том, что ацетилхолин подвергается ферментативному гидролизу. Второй путь – это энергозависимый активный транспорт ацетилхолина в нейрон, где он накапливается для последующего повторного использования.
В адренорецепторах существует два вида рецепторов для норадреналина: α- и β-адренергические рецепторы. Эти рецепторы можно отличить друг от друга по специфическим реакциям, которые они вызывают, а также по тем специфическим агентам, которые способны блокировать данные реакции.
β-Адренергические рецепторы включают эфферентную клетку с по-
мощью аденозин-3',5'-монофосфата, или цАМФ – универсального «второго посредника» между гормонами и различными функциями клеток, на которые воздействуют гормоны.
Установлено, что как только β-адренергический рецептор, расположенный на наружной поверхности мембраны эффекторной клетки, начинает взаимодействовать с норадреналином, на внутренней поверхности клеточной мембраны активируется фермент аденилатциклаза. Затем в клетке аденилатциклаза превращает АТФ в цАМФ; последний в свою очередь способен оказывать влияние на метаболизм клетки. Этот сложный ряд последовательных реакций может быть заблокирован пропранололом – веществом, препятствующим связыванию норадреналина с β-адренергическим рецептором.
Катехоламины
В метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (—NH2) у норадреналина, серотонина, дофамина и адреналина, тем самым инактивируя указанные медиаторы.
Адренергическая и холинергическая системы головного мозга тесно взаимодействуют с другими системами мозга, в частности использующими серотонин в качестве медиатора. В основном серотонинсодержащие нейроны сосредоточены в ядрах мозгового ствола. Нейромедиаторная роль серотонина осуществляется в результате взаимодействия серотонина со специфическими серотонинергическими рецепторами.
Ацетилхолин -представляет собой сложный эфир уксусной кислоты и холина. Он синтезируется в нервной клетке из холина и активной формы ацетата – ацетилкоэнзима. Хранится ацетилхолин в синаптических везикулах нервных окончаний и высвобождается путем экзоцитоза. Сигналом для высвобождения ацетилхолина является увеличение концентрации ионов Са2+ в нервных окончаниях. Расщепление медиатора в синаптической щели производится ферментом ацетилхолинэстеразой.
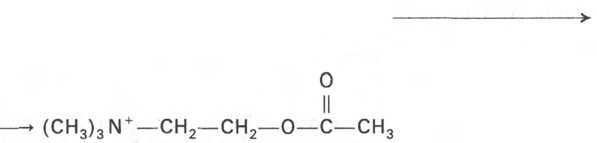
Ацетилхолин
Катехоламины синтезируются в нервной системе из тирозина в несколько этапов:
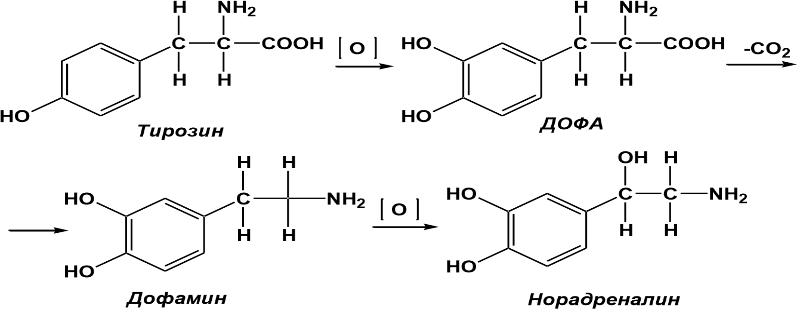
Норадреналин – принимает участие в процессах узнавания и запоминания, а также в адаптивных процессах. Развитие депрессивных состояний связывают также с уменьшением норадренергической активности в ЦНС.
Серотонин – является производным 5-гидрокситриптофана, принимает участие в регуляции таких психоэмоциональных состояний как: тревога, агрессия, сексуальное поведение, в контроле циклов физиологического сна.
Нарушение обмена серотонина и работы серотониновых рецепторов способствуют развитию депрессивных состояний, шизофрении, алкоголизма, наркомании.
Биогенные амины синтезируются из предшественников с помощью ферментов декарбоксилаз, а обезвреживаются за счет аминооксидаз (моноаминооксидазы, диаминооксидазы).
Глицин- тормозной медиатор, по вызываемым эффектам подобен гамк.
Гистамин(он же β-имидазолилэтиламин) – биогенный амин, нейромедиатор. Помимо участия в аллергических реакциях, его функции в организме также включают в себя регуляцию суточного ритма, тонуса сосудов и активности секреторных клеток слизистой оболочки. Даже половое влечение отчасти зависит от гистаминэргических систем.
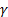 -аминомасляная кислота
-аминомасляная кислота
Важным нейромедиатором, выполняющим тормозные функции, является γ-аминомасляная кислота (ГАМК), количество которой в головном мозге во много раз больше, чем других нейромедиаторов. ГАМК увеличивает проницаемость постсинаптических мембран для ионов К+ и тем самым отдаляет мембранный потенциал от порогового уровня, при котором возникает потенциал действия; таким образом, ГАМК – это тормозной нейромедиатор. ГАМК образуется при декарбоксилировании глутамата в реакции, катализируемой 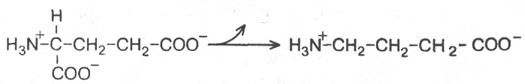
глутаматдекарбоксилазой:
Глутамат γ-Аминобутират
Нейропептиды
Пептиды
Имеют в своем составе от трех до нескольких десятков аминокислотных остатков. Функциониуют только в высших отделах нервной системы.
Эти пептиды, как и катехоламины, выполняют функцию нейромедиаторов и гормонов. Передают информацию от клетки к клетке по системе циркуляции. Сюда относятся:
а) нейрогипофизарные гормоны (вазопрессин, либерины, статины). Эти вещества одновременно и гормоны и медиаторы;
б) гастроинтестинальные пептиды (гастрин, холецистокинин). Гастрин вызывает чувство голода, холецистокинин вызывает чувство насыщения, а также стимулирует сокращение желчного пузыря и функцию поджелудочной железы;
в) опиатоподобные пептиды (или пептиды обезболивания). Образуются путем реакций ограниченного протеолиза белка-предшественника проопиокортина. Взаимодействуют с теми же рецепторами, что и опиаты (например, морфин), тем самым имитируют их действие. Общее название - эндорфины - вызывают обезболивание. Они легко разрушаются протеиназами, поэтому их фармакологический эффект незначителен;
г) пептиды сна. Их молекулярная природа не установлена. Известно лишь, что их введение животным вызывает сон;
д) пептиды памяти (скотофобин). Накапливается в мозге крыс при тренировке на избегание темноты;
е) пептиды - компоненты ренин-ангиотензиновой системы. Показано, что введение ангиотензина-II в центр жажды головного мозга вызывает появление этого ощущения и стимулирует секрецию антидиуретического гормона.
Образование пептидов происходит в результате реакций ограниченного протеолиза, разрушаются также под действием протеиназ.
Механизмы памяти
Память не сосредоточена в одном строго локализованном участке мозга, подобно центрам зрения, слуха, речи и т.д. В то же время память – не свойство всего мозга в целом. Субстратом памяти человека являются нейроны.
Память человека нельзя рассматривать в отрыве от его деятельности, так как не познание познает, не мышление мыслит, не память запоминает и воспроизводит, а познает, мыслит, запоминает и воспроизводит человек, определенная личность.
В последние годы отчетливо показано, что обучение животного новым навыкам отражается на химизме мозговых клеток (нейронов): меняются количество уридина в цитоплазматической РНК, степень метилирования ДНК и фосфорилирования ядерных белков. Применение стимуляторов и веществ – предшественников РНК облегчает обучение, а введение блокаторов синтеза РНК, напротив, затрудняет этот процесс. Существуют данные, что после запоминания информации меняется антигенный состав мозговой ткани. Принято выделять несколько форм биологической памяти: генетическую, иммунологическую и нейрологическую. Биохимические основы генетической памяти более или менее ясны. Ее носителем является ДНК клетки. Следующей по сложности формой памяти является иммунологическая память. Этот вид памяти хотя и включает элементы генетической памяти, но находится на более высокой ступени сложности. Наконец, система нейрологической памяти еще более сложна. Эта форма в свою очередь может быть разделена на кратковременную память (КП) и долговременную память (ДП). В основе КП, по всей вероятности, лежит «циркуляция» информации, полученной в виде импульсов, по замкнутым цепям нейронов. При этом синаптический эффект, изменения ядерно-ядрышкового аппарата, выброс в цитоплазму нейрона биологически активных веществ и сопутствующая этим процессам перестройка обмена веществ клетки – все это может расцениваться как показатели функционирования КП.
Включение блоков ДП обеспечивается примерно через 10 мин после прихода информации в клетку. За это время происходит перестройка биологических свойств нервной клетки. Ряд исследователей считают, что афферентная импульсация, приходящая в нервные клетки во время обучения, вызывает либо количественную активацию синтеза РНК и белка, что может приводить к установлению новых синаптических связей и перестройке существующих, либо наступающая активация синтеза нуклеиновых кислот и белка носит целенаправленный, специфический характер, а синтезированные молекулы являются хранилищем информации.
Роль нейромедиаторов в регуляции памяти. Процессы памяти тесно связаны с модификацией синтетических процессов. Поэтому химические передатчики нервного возбуждения должны играть в этом принципиальную роль. Накоплен большой эспериментальный материал о значении нейромедиаторов в процессах памяти и обучения. Полученные к настоящему времени результаты свидетельствуют о большой значимости основных медиаторов (ацетилхолин, норадреналин, дофамин, серотонин, ГАМК) в этих процессах, хотя конкретные формы участия каждого медиатора зависят от того, какой именно тип информации запоминается. Например, показано, что снижение содержания ацетилхолина в мозге ингибиторами холинацетилазы нарушает обучение, а его повышение ускоряет выработку оборонительных навыков. Серотонин облегчает выработку и хранение навыков, основанных на положительном (пищевом) подкреплении, и отрицательно влияет на формирование оборонительных реакций
Олигопептиды - регуляторы памяти. Установлено, что некоторые олигопептиды, представляющие собой молекулы из небольшого числа аминокислотных остатков, способны модифицировать процесс обучения и влиять на степень выработки, хранения и угасания приобретенных поведенческих реакций. Из пептидов, относящихся к числу гормонов, наиболее выраженное действие на процессы обучения и памяти оказывают гормоны гипофиза – адренокортикотропный гормон (АКТГ) и вазопрессин. При изучении влияния АКТГ на процессы памяти было показано, что главная роль в его действии принадлежит фрагменту АКТГ 4 - 1 0 , который оказывает на эти процессы практически такой же эффект, как и целый гормон.






