

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Топ:
Характеристика АТП и сварочно-жестяницкого участка: Транспорт в настоящее время является одной из важнейших отраслей народного...
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Характеристика АТП и сварочно-жестяницкого участка: Транспорт в настоящее время является одной из важнейших отраслей народного хозяйства...
Интересное:
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Аура как энергетическое поле: многослойную ауру человека можно представить себе подобным...
Уполаживание и террасирование склонов: Если глубина оврага более 5 м необходимо устройство берм. Варианты использования оврагов для градостроительных целей...
Дисциплины:
|
из
5.00
|
Заказать работу |
Содержание книги
Поиск на нашем сайте
|
|
|
|
Представим, что мы являемся совершенными «машинами Корбаллиса» – идеально симметричными организмами. Наше левое полушарие – точное зеркальное отражение правого, включая нейроны, связи и прочие особенности. Как бы мы себя вели? Смогли бы мы по‑прежнему перемещаться в пространстве, отличать левую сторону от правой или называть зрительные объекты? Ошибки какого типа мы бы совершали? Были бы они похожи на зеркальные ошибки маленьких детей?
Здесь мы должны отказаться от одного ошибочного представления. Даже если наш организм был бы абсолютно симметричным, вплоть до самых глубоких тайников его нервной системы, то он не обязательно путал бы лево и право во всех ситуациях без исключения. На самом деле такой организм смог бы выполнять пространственно ориентированные действия в ответ на асимметричную стимуляцию. Вообразите, например, что он видит тигра, приближающегося справа. Допустим, он заранее запрограммирован на то, чтобы при появлении хищника справа бежать влево. Симметрия его тела подразумевает, что при появлении тигра слева он должен предпринять симметричное действие (бежать вправо). Следовательно, любой наблюдатель неизбежно придет к заключению, что этот совершенно симметричный организм превосходно отличает левую сторону от правой.
Наш гипотетический симметричный организм одинаково откликается на два зеркальных отражения только в том случае, когда реакция носит условный и непространственный характер. Если он научится произносить слово «тигр» при виде правой стороны тигра, то симметрия его тела заставит его дать аналогичный ответ и при появлении левой стороны животного. Другими словами, он может распознать и даже назвать тигра независимо от его ориентации в пространстве. Эта полезная особенность проявляется сразу же, как только организм видит хищника под новым углом.
Обладание симметричным мозгом и поддержание этой особенности в процессе научения имеет два преимущества:
• Симметрия тела позволяет организму определять зрительные объекты инвариантно, независимо от их левосторонней или правосторонней ориентации в пространстве.
• Это не мешает совершению надлежащих движений и жестов, ориентированных в пространстве.
Существует, однако, два вида операций, которые симметричному организму не под силу. Первый из них – это произвольная реакция на асимметричный объект, но не на его зеркальное отражение. В повседневной жизни без таких операций трудно обойтись. Например, мы произносим как [би] букву «b», но не «d», и говорим «левая» при виде левой руки, но не правой. Идеально симметричный организм не смог бы выполнить эти задачи, какими бы простыми они ни казались.
Вторая операция, недоступная зеркальному мозгу, состоит в выполнении асимметричных жестов в ответ на команду, не содержащую пространственной информации. Предположим, вас просят поднять только правую руку по сигналу «правая». Идеально симметричный организм не сможет этого сделать: связи, соединяющие слуховые области с моторными командами, вызовут движение не только правой, но и левой руки. Следовательно, такой организм будет выполнять жесты сразу двумя конечностями или, при наличии некоторого шума, беспорядочно поднимать то одну, то другую.
Удивительно, но теоретические ограничения симметричного организма в точности совпадают с экспериментальными наблюдениями за животными и маленькими детьми. Ни один ребенок не испытывает трудностей с называнием знакомого предмета, даже если смотрит на него под новым углом. Бросая мяч, ни один малыш не станет поднимать левую руку, если держит мяч в правой. С другой стороны, все дети не только с трудом различают буквы «b» и «d», но и регулярно путают левую и правую руки.
Таким образом, хотя концепция симметричного мозга, активно поддерживающего свою пропорциональность в процессе научения, представляется несколько надуманной, она заслуживает самого пристального внимания, поскольку отражает некоторые когнитивные ограничения, присущие человеку в совладании с левым и правым. Если так, нам нужны конкретные доказательства того, что межполушарный перенос действительно навязывает лево‑правую инверсию, постулируемую Корбаллисом и Билом. На самом деле все вышеизложенное не настолько оторвано от биологии, как может показаться на первый взгляд. Результаты нескольких экспериментов свидетельствуют о тесной связи восприятия симметрии с самой симметрией мозга.
Симметрия одного нейрона
Нейрофизиологи только начинают изучать нейрональный базис восприятия симметрии, но имеющиеся данные уже указывают на зеркальную симметрию как на один из основных инвариантов, извлекаемых нейронами нижней височной коры. Именно там расположена «буквенная касса» мозга. Как я уже отмечал ранее, нейроны нижней височной коры, независимо от изменения угла обзора, склонны отдавать предпочтение определенным зрительным объектам. Нейрон, который предпочитает, скажем, лицо Дженнифер Энистон, сохраняет свой выбор безотносительно к его размеру и положению на сетчатке. Недавно нейрофизиологи сообщили, что некоторым из этих клеток свойственна инвариантность к зеркальной симметрии[447]: профиль их отклика остается идентичным даже при лево‑правой инверсии изображения.
Важно отметить, что эти нейроны часто демонстрируют большую инвариантность относительно горизонтальной инверсии, нежели вертикальной. Например, нейрон, который интенсивно реагирует на букву «р», точно так же будет отвечать на ее зеркальное отражение по оси «лево – право» («q»), но не на буквы, симметричные по вертикальной оси. Другими словами, этот нейрон отличает «р» от «b», но не «р» от «q».
В совокупности нейроны нижней височной коры реализуют перцептивную инвариантность: они позволяют распознавать объект независимо от его размера и положения в пространстве. Электрофизиологические данные подразумевают, что противопоставление «лево – право» входит в число несущественных различий, которые наша вентральная зрительная система систематически игнорирует.
Если теория Корбаллиса и Била верна, специально учиться зеркальной инвариантности не нужно. Она принадлежит к структурным инвариантам, изначально присущим организации наших межполушарных связей. Исследования нейрофизиолога Никоса Логотетиса[448] убедительно подтверждают эту гипотезу. Логотетис обучил макак распознавать необычные трехмерные фигуры, похожие на скрученные скрепки для бумаг (рис. 7.4). Он хотел узнать, смогут ли обезьяны определить их позже, когда увидят предметы с другой стороны. Во время обучения Логотетис тщательно следил за тем, чтобы животные видели каждый объект только под одним углом – ощущение глубины создавало лишь минимальное вращение вокруг вертикальной оси. Но было ли этого достаточно для инвариантного распознавания? Чтобы ответить на этот вопрос, Логотетис показывал макакам различные ракурсы объекта одновременно с другими отвлекающими фигурами.
Оказалось, что обезьяна не могла распознать предмет, когда он был представлен с отклонением от усвоенной ориентации на 40 градусов и более. Однако, если его поворачивали на 180 градусов, показатели распознавания резко возрастали. Аналогично вели себя и нейроны нижней височной коры: большинство клеток энергично срабатывали при предъявлении объекта в усвоенной ориентации, переставали реагировать, когда его поворачивали на 40 или 50 градусов, и вновь активировались при достижении угла 180 градусов. Почему именно 180 градусов? Если повернуть проволочный объект на 180 градусов, он будет выглядеть почти так же, как зеркальное отражение исходного объекта. Скрутите скрепку в букву «b», поверните ее на 180 градусов и получите «d».

Рис. 7.4. Усвоив определенную форму, зрительная система приматов обобщает результаты научения на ее зеркальное отражение. В этом эксперименте обезьяну научили распознавать проволочный объект, представленный под определенным углом. Затем ей показали тот же объект, но под разными ракурсами (верхний график). Животное распознавало его при предъявлении под усвоенным углом, а также при повороте на 180 градусов, что соответствовало зеркальному отражению исходного вида. После обучения нейроны реагировали как на первоначальную форму, так и на ее зеркальное отражение (нижний график) (по материалам статьи Logothetis, Pauls, & Poggio, 1995). Используется с разрешения Oxford University Press.
Вкратце, нейроны в нижней височной коре могут распознавать зеркальное отражение усвоенной фигуры, даже когда видят его впервые. Никакого дополнительного обучения для этого не требуется. Изучение нейронов, преимущественно реагирующих на лица, подтверждает этот вывод. Многие клетки одинаково активно срабатывают на два зеркально‑симметричных вида одного лица, например на левый и правый профили[449]. Удивительно, но такая инвариантность присутствует даже у новорожденных детенышей обезьян[450]. Что касается человека, то младенцы могут распознавать зеркальное отражение знакомого предмета уже в 4 месяца[451]. Похоже, все мы с рождения обладаем поразительной чувствительностью к подобной симметрии.
Исследования Логотетиса подчеркивают любопытный контраст между вращением и симметрией. Инвариантность относительно вращения, по всей видимости, не входит в число исходных возможностей нашей зрительной системы. Иными словами, чтобы распознавать объект под любым углом, мы должны знать, как он выглядит при повороте. Очевидно, на этой стадии наша зрительная система не обладает глубокими знаниями трехмерных форм и опирается на двумерные представления, которые видела ранее. Симметрия этому правилу не подчиняется.
Нейровизуализационные эксперименты с участием макак и людей показали удивительную чувствительность к симметрии, свойственную зрительной коре на ранней стадии. Представьте совокупность хаотично расставленных точек. При добавлении всего нескольких симметрично расположенных точек активность латеральных областей затылочно‑височной коры резко возрастает. Способность обнаруживать симметрию напрямую связана с повышенной активностью нейронов[452]. Хотя эволюция не готовила нас к такой задаче, эти точки резонируют в наших зрительных цепях, мгновенно срабатывающих при любом намеке на симметрию.
Симметричные связи
На сегодняшний день геометрии нейронных связей, лежащих в основе восприятия симметрии, было посвящено не так много исследований. Возможно, Бил и Корбаллис правы, и ключ к восприятию симметрии – это мозолистое тело с его многочисленными симметричными связями, соединяющими два полушария. Однако в отсутствие точных данных нам не остается ничего другого, кроме как строить догадки, опираясь на разрозненные, но любопытные факты. Я уже рассказывал о новаторских экспериментах Павлова и Била на собаках и голубях. Оба вида с трудом различают пары зеркальных изображений. Голубь, например, легко усваивает разницу между вертикальными и горизонтальными палочками (– или |), но показывает плохие результаты, когда они поворачиваются на 45 градусов (/ или \). Как ни странно, рассечение мозолистого тела фактически облегчает задачу. Это свидетельствует о том, что межполушарные связи действительно вносят свой вклад в восприятие симметрии.
Но какую именно роль они играют? Другие эксперименты показывают, что перенос информации из одного полушария в другое через мозолистое тело переворачивает зрительное пространство по подобию перчатки. В результате левая и правая стороны меняются местами. В 1960‑х годах Нэнси Мелло из Гарварда опубликовала серию статей о чувстве симметрии у голубей[453]. Этим птицам присуща одна очень полезная анатомическая особенность: в отличие от млекопитающих, правый глаз голубя проецирует изображение исключительно на зрительные области левого полушария, а левый глаз – на зрительные области правого. Это означает, что если закрыть голубю один глаз, то визуальная информация будет поступать только в одно полушарие[454].
Мелло научила голубей различать одним глазом два зрительных образа, например Λ и V или < и >. Птицы получали вознаграждение, если клевали определенную из фигур. Когда связь была установлена, Мелло решила выяснить, как она переносилась на другой глаз. Сюрприз! Голуби показывали хорошие результаты с вертикальной симметрией (Λ или V), но систематически терпели неудачу с симметрией «лево – право» (< или >). Голубь, обученный клевать форму <, видимую правым глазом, систематически клевал симметричную фигуру >, воспринимаемую левым глазом. Таким образом, усвоенная зрительная информация действительно передавалась из одного полушария в другое, но при этом левая и правая стороны менялись местами, как и предполагает модель Корбаллиса и Била.
Годом позже аналогичные эксперименты были проведены на животных, более близких к человеку, – макаках. Джон Нобл из Университетского колледжа Лондона сначала рассекал оптическую хиазму, чтобы каждый глаз проецировал только на противоположное полушарие, как у голубей. Затем он обучал животных различать два симметричных объекта одним глазом, после чего проверял обобщение на другой глаз. Нобл наблюдал то же, что и Мелло: загадочный перенос зеркальных изображений. Подобно голубям, обезьяны систематически выбирали зеркальное отражение фигуры, которой их обучали.
В ходе дальнейших экспериментов Нобл показал, что перенос зрительных знаний осуществляется либо через мозолистое тело, либо через другой пучок волокон, так называемую переднюю комиссуру. Зеркальный перенос исчезал, если оба пучка были рассечены, но сохранялся, если один из них оставался нетронутым. В одном эксперименте Нобл удалил межполушарные связи после периода обучения, но до проверки обобщения на другой глаз. Зеркальная инверсия наблюдалась по‑прежнему. Это наводит на мысль, что мозолистое тело было задействовано в период обучения, но не во время тестирования.
Нобл пришел к выводу, что у обезьяны каждый учебный эпизод сопровождается зеркальным научением в другом полушарии. Этот механизм мог бы объяснять особые трудности, которые животные и люди испытывают при различении левой и правой сторон. При бинокулярном зрении зеркальное научение гарантирует, что оба полушария получают как первичный зрительный сигнал, так и вторичное зеркальное отражение внешнего мира. Если наблюдаемая сцена симметрична, обе репрезентации идентичны и усиливают друг друга. Однако если сигналы асимметричны, например «b» и «d», два внутренних представления конкурируют друг с другом. Этот конфликт замедляет научение, даже если в итоге побеждает первичная репрезентация.
Если эти рассуждения верны, анатомией межполушарных связей управляет принцип симметрии. Иначе говоря, симметричные секторы зрительных областей должны быть связаны напрямую, проекциями один к одному. Так оно и есть[455]. Это является еще одним аргументом в пользу модели Корбаллиса и Била. Геометрия межполушарных связей так же безупречна, как крылья бабочки. Аксон нейрона, расположенного в левой зрительной коре, сначала идет перпендикулярно поверхности коры, после чего присоединяется к пучку волокон мозолистого тела. Достигнув противоположного полушария, он направляется к месту, точно симметричному исходной точке (рис. 7.5). Такие нейроны были обнаружены в первичной зрительной коре крыс и мышей, а также во вторичной зрительной области макак. Их функция неизвестна, но их геометрия идеально подходит для отзеркаливания зрительных сигналов, что существенно облегчает обнаружение симметричных объектов.
Если этими симметричными связями иногда пренебрегают, то только потому, что они составляют лишь небольшой процент контактов, соединяющих два полушария. Главный пучок межполушарных волокон не симметричен. Он служит для «сшивания» двух корковых репрезентаций зрительного мира и дает ответ на известную загадку: почему мы воспринимаем один целостный зрительный мир, когда объективно он разделен на две половины, которые кодируются разными полушариями (см. рис. 7.5)? Межполушарные связи играют важную роль в восстановлении функционального единства мира, несмотря на анатомическое разграничение. «Сшивающие» связи концентрируются в области разрыва (вертикальной срединной линии) и соединяют секторы левого и правого полушарий, кодирующие одинаковые зрительные локации.
Исследования Хайме Олаварриа и его коллег из Сиэтла позволили выявить конкуренцию, которая возникает в процессе развития между принципом зрительной целостности, реализуемым «сшивающими» связями, объединяющими два полушария, и принципом симметрии, требующим симметричных проекций большего масштаба[456]. Многочисленные симметричные связи, присутствующие при рождении, вероятно, заложены градиентами химического притяжения. Однако уже в первые дни жизни они подвергаются активному прунингу. Благодаря этому процессу «сшивающие» связи, обеспечивающие целостность зрительного поля, в конечном итоге преобладают.
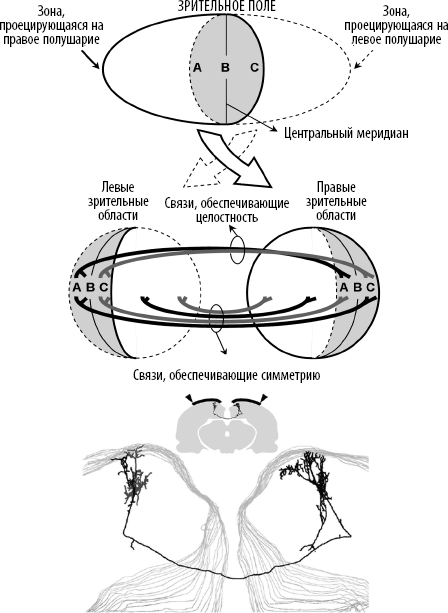
Рис. 7.5. Целостность и зеркальная симметрия – основные принципы, управляющие архитектурой зрительных связей между двумя полушариями. Верхний рисунок иллюстрирует хорошо известный факт, что левая половина зрительного поля проецируется на правое полушарие, а правая половина – на левое, всего лишь с незначительным перекрытием в центре (по материалам статьи Olavarria & Hiroi, 2003). Если мы не воспринимаем никаких разрывов в зрительной сцене, то это потому, что межполушарные связи обеспечивают функциональную целостность зрительного поля, соединяя области коры, кодирующие одну и ту же точку. Эти «сшивающие» связи касаются только центральной области сетчатки и не являются симметричными. Существуют и другие межполушарные связи, которые соединяют симметричные точки зрительного поля даже на периферии. Вероятно, они играют особую роль в восприятии симметрии и зеркальном обобщении. На нижнем рисунке мы видим их безупречную геометрию: один нейрон в первичной зрительной коре крысы проецируется на симметричные области левого и правого полушарий (по материалам статьи Houzel, Carvalho, & Lent, 2002). Использовано с разрешения Brazilian Journal of Medical and Biological Research.
Как ни странно, прунинг происходит даже в отсутствие контакта с окружающей средой. Еще в то время, когда плод находится внутри утробы матери, спонтанные волны нейронной активности, проходящие через сетчатку, сообщают нейронам в левом и правом полушариях, что они кодируют одну и ту же ретинальную локацию. У крыс «сшивающие» связи формируются за несколько дней до того, как они впервые откроют глаза. Нарушение самопроизвольной активности сетчатки ведет к их дезорганизации. В этом случае сшивающие аксоны проигрывают гонку, и в мозге наблюдается избыток симметричных связей.
Я столь подробно остановился на этих биологических особенностях только потому, что все они могут играть важную роль в развитии дислексии. В частности, они предполагают, что сбои в процессе синаптического прунинга в последние месяцы беременности или первые месяцы жизни могут привести к переизбытку симметричных зрительных связей у ребенка. Не потому ли некоторым детям так трудно различать лево и право или букву «b» от «d»? Прежде чем подробно обсудить этот вопрос, давайте посмотрим, каким образом мозг проводит такие различия.
Спящая симметрия
Если бы наша зрительная система делала все симметричным, то мы бы постоянно совершали зеркальные ошибки – например, мы бы не могли отличить «р» от «q» и левый ботинок от правого. К счастью, часть нашей зрительной системы сохраняет способность различать левую и правую стороны.
Как указывают ряд авторов[457], наша зрительная кора разделена на два основных функциональных потока, как будто в нашем мозге существует две отдельных зрительных системы. Вентральный путь, о котором я говорил ранее, отвечает за инвариантное распознавание объектов. Он крайне чувствителен к характеру изображения, форме и цвету, но игнорирует размер и пространственную ориентацию – в том числе левое и правое. Второй зрительный путь – дорсальный – проходит через теменную кору и в первую очередь связан с пространством и действием. В этом маршруте цвет и точная природа объекта не так важны, как расстояние, положение, скорость и ориентация в пространстве. Все эти параметры помогают определить, как нам следует взаимодействовать с этим объектом. Дэвид Милнер и Мел Гудейл кратко подытоживают это фундаментальное различие: вентральному маршруту важно «что», дорсальному – «как».
«В начале было дело!» – утверждает Фауст Гете. Умению отличать левую сторону от правой мы, по всей вероятности, обязаны дорсальному пути. В детстве мы узнаем, что правой рукой рисуют и пишут. В римско‑католических семьях правой рукой крестятся. Система «как» должна уметь отличать правую сторону от левой, потому что эта информация имеет решающее значение для жестов. Мы выполняем разные движения, чтобы взять кастрюлю, ручка которой направлена влево или вправо от нас: дорсальный путь автоматически адаптирует наши действия к ориентации объекта, даже если вентральная система, которая нечувствительна к расположению, говорит нам, что в обоих случаях кастрюля одна и та же. Кроме того, именно дорсальная система позволяет нам представлять сугубо виртуальные жесты или движения. Мы полагаемся на нее всякий раз, когда мысленно поворачиваем объекты, читаем карту или пытаемся определить, что изображено на картинке: два отличных друг от друга объекта или один и тот же, но под разными углами.
Вкратце, наш мозг ведет себя так, как если бы в нем жили два маленьких человечка. Первый, которого я буду называть коллекционером, распознает и помечает объекты, но почти не обращает внимания на их расположение и ориентацию в пространстве. Второй, «рабочий», действует, сравнивает, манипулирует… но не может определять объекты, которые попадают ему в руки.
Обычно две зрительные системы взаимодействуют настолько тесно, что мы даже не подозреваем о таком внутреннем разделении труда. Оно становится явным только при некоторых повреждениях мозга. Шотландский исследователь Оливер Тернбулл обследовал десятки больных с различными травмами мозга, которые привели к диссоциации между двумя зрительными системами. Возьмем двух пациентов: Д. М., пережившего черепно‑мозговую травму, и Л. Дж., перенесшую множественные инсульты в правой височно‑теменной области[458]. Тернбулл показывал им изображения знакомых предметов, ориентированных в разных направлениях: перевернутый вертолет, стоящий вертикально кенгуру, лежащий телефон и так далее. Д. М. не мог их назвать, а Л. Дж. справлялась с задачей весьма неплохо. Когда их спрашивали, изображены ли предметы в правильной ориентации, происходило обратное: Д. М., который не узнавал объекты, отвечал безупречно, в то время как Л. Дж. систематически терпела неудачу. У Д. M. был поврежден вентральный путь «что», а у Л. Дж. больше не функционировал дорсальный путь «как».
Аналогичная диссоциация между идентичностью объекта и ориентацией наблюдается и у макак. Повреждения вентральной височной коры лишают животное способности различать такие простые фигуры, как круг и крест, но не оказывают никакого влияния на умение определять ориентацию (например, «6» и «9» или «b» и «d»). После повреждения дорсального теменного маршрута происходит обратное: животное, подобно маленькому ребенку, может отличить «а» от «b», но не «b» от «d»[459].
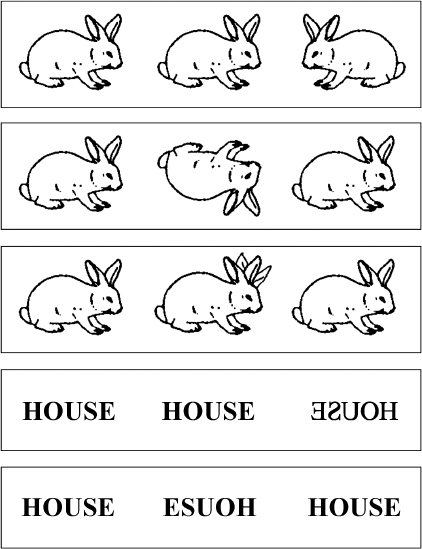
Рис. 7.6. После повреждения мозга некоторые пациенты больше не могут различать зеркальные изображения. Чтобы исследовать их восприятие пространственной ориентации, Оливер Тернбулл и Розалин Маккарти (1996) разработали специальный тест: они предлагали испытуемым три почти одинаковых изображения и просили выбрать то, которое отличается от двух других. Пациент Р. Дж. систематически терпел неудачу, когда разница заключалась в направленности «лево – право» (верхний ряд), но преуспевал во всех других испытаниях, включая задания на ориентацию слов и букв (нижние ряды). Используется с разрешения Neuropsychologia.
Дорсальный путь «как» включает множество цепей, организацию которых мы только начинаем понимать. Одни области связаны с движениями кистей рук, другие – с программированием движений глаз, третьи – с кодированием разных типов пространственных отношений. Параметры расстояния, размера, количества, формы, движения и ориентации задействуют различные подобласти теменной коры, пересекающиеся лишь частично. Например, после повреждения правой теменной области многие пациенты частично теряют способность различать лево и право. Этот тип поражения обнажает дремлющую симметрию, вычисляемую вентральной затылочно‑височной системой «что».
Оливер Тернбулл и Розалин Маккарти описали один из первых случаев этого синдрома «зеркальной слепоты». Пациент Р. Дж. (61 год), у которого было диагностировано двустороннее поражение теменной области вследствие коронарного тромбоза[460], настаивал, что не видит никакой разницы между объектом и его отражением. Если два изображения накладывали друг на друга, он признавал, что картинки совпадают не идеально, но как только их разделяли, он переставал их различать. «Я понимаю, что они должны быть разными, – говорил он, – но когда я смотрю на одну, а потом на другую, они кажутся мне абсолютно одинаковыми».
Чтобы изучить эту странную патологию, Тернбулл и Маккарти показывали Р. Дж. три почти идентичных изображения и просили выбрать одно лишнее. Р. Дж. с легкостью замечал даже незначительные изменения в идентичности объекта, например кролика с четырьмя ушами или перевернутого вниз головой. Но он не мог обнаружить кролика, смотрящего вправо, стоящего в одном в ряду сородичей, которые смотрели влево (рис. 7.6). С тех пор были зарегистрированы и другие случаи «зеркальной слепоты»[461]. Все эти пациенты были неспособны отличить объект от его зеркального отражения, будь то рисунки животных, инструменты или незнакомые проволочные объекты. Повреждение дорсально‑теменного пути лишило их информации о левой и правой сторонах, а здоровая вентральная зрительная система слепа к ориентации.
Разбитое зеркало
Как ни странно, существует одно примечательное исключение в случае зеркальной слепоты, которое возвращает нас к сути предмета нашего исследования – к чтению. Во время чтения пациенты с зеркальной слепотой не испытывают никаких трудностей с различением левой и правой сторон. Всем им удается верно определить, как написаны буквы и слова: нормально или в зеркальном отражении. Они легко обнаруживают лишнее слово, причем даже тогда, когда перевернуты отдельные буквы (РАЗУМ – 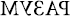 ), отражено все слово (УЛЫБКА – АКБЫЛУ) или и то, и другое (ЗРЕНИЕ –
), отражено все слово (УЛЫБКА – АКБЫЛУ) или и то, и другое (ЗРЕНИЕ –  ). Все эти различия так же очевидны для них, как и для здоровых людей, хотя из‑за повреждения мозга они не видят никакой разницы между невербальными зеркальными формами, например
). Все эти различия так же очевидны для них, как и для здоровых людей, хотя из‑за повреждения мозга они не видят никакой разницы между невербальными зеркальными формами, например  и
и  .
.
Это наблюдение подтверждает, что чтение подчиняется особым правилам. У опытных читателей часть вентрального пути отучается обобщать зеркальные образы. На ранних стадиях формирования навыка чтения мы задействуем дорсальную систему, чтобы различать такие буквы, как «b» и «d». Только она может определить, куда «указывает» буква: влево или вправо. Вентральная система говорит нам, что оба изображения представляют одну и ту же форму. Постепенно она учится игнорировать симметрию. Она перестает рассматривать «b» и «d» как два вида одного объекта и «поручает» их отдельным популяциям нейронов, которые не обобщают зеркальные инверсии. Эти детекторы асимметричных букв служат отправными точками для извлечения подробной статистики о чтении, которая применима только к правильно ориентированным словам. В результате затылочно‑височная кора приобретает асимметричную нейрональную иерархию для зрительного распознавания слов. В отличие от соседних областей коры, участвующих в идентификации объектов и лиц и продолжающих обобщать лево‑правые инверсии, архитектура, отвечающая за чтение, не смешивает зеркальные отражения.
Подобный процесс нарушения симметрии, по всей вероятности, затрагивает только письменные знаки. Разумеется, он принесет пользу лишь в том случае, если ассиметрична сама система письма. А это не всегда так. Египетские иероглифы, например, писались в обоих направлениях: справа налево и слева направо. В последнем случае все символы были перевернуты. Направление чтения диктовали фигуры человека и животных: их головы всегда были повернуты к началу строки. То же самое можно сказать и о древнегреческом языке, для которого был характерен бустрофедон – способ письма, название которого буквально означает «как бык пашет»: одна строка писалась слева направо, а следующая – справа налево, в зеркальном отражении. Таким образом, ни египтянам, ни грекам было не нужно переучивать вентральную систему. На самом деле такая двунаправленная письменность никогда бы не была принята, если бы человеческая зрительная система не воспринимала формы  и
и 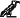 как идентичные. По сей день некоторые системы письменности не требуют нарушения симметрии. В частности, в китайском языке существует всего несколько зеркальных символов, сравнимых с нашими «b» и «d», например
как идентичные. По сей день некоторые системы письменности не требуют нарушения симметрии. В частности, в китайском языке существует всего несколько зеркальных символов, сравнимых с нашими «b» и «d», например  и
и  или
или  и
и  . Другие системы, например тамильское письмо, полагаются исключительно на такие изящные знаки, как
. Другие системы, например тамильское письмо, полагаются исключительно на такие изящные знаки, как 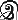 ,
,  или
или 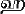 , заведомо исключающие зеркальную путаницу. Подобные культурные различия приводят к снижению перцептивной компетенции: носители тамильского языка легко путают зеркальные отражения. Например, они уверены, что в фигуре
, заведомо исключающие зеркальную путаницу. Подобные культурные различия приводят к снижению перцептивной компетенции: носители тамильского языка легко путают зеркальные отражения. Например, они уверены, что в фигуре  присутствует треугольник
присутствует треугольник  [462].
[462].
Нарушенная симметрия… Или скрытая симметрия?
Что можно сказать о носителях европейских языков, которые читают латинский алфавит и могут отличить «b» от «d»? У них восприятие симметрии исчезает полностью? Пока неизвестно. На сегодняшний день мы не знаем, действительно ли наша зрительная система перестает обобщать зеркальные образы слов. Может, мы просто этого не осознаем? Сэмюэл Ортон полагал, что правое полушарие переворачивает буквы даже у опытного читателя. По мнению Ортона, обучение чтению сводилось к сосредоточению на правильной ориентации слов. Их зеркальное отражение, хотя и оставалось без внимания, все же присутствовало в недоминантном полушарии. Зеркало было не разбито, а просто спрятано.
Мысль о том, что нас бессознательно преследуют зеркальные отражения прочитанных слов, может быть не столь абсурдна, как кажется на первый взгляд. По всей вероятности, на начальных стадиях зрительного анализа буквы представляют собой бессмысленные фигуры, которые обрабатываются так же, как и любой другой визуальный образ. Если это так, в результате межполушарного переноса они должны переворачиваться по подобию перчатки. И действительно, столкнувшись с зеркальным отражением слова, мы можем расшифровывать его побуквенно. Хотя это требует определенных усилий, очевидно, что идентификация отдельных букв возможна даже в отсутствие доступа к более глубинной системе, позволяющей обрабатывать зрительную форму всего слова.
Многочисленные примеры из жизни свидетельствует о том, что на моторном уровне все мы наделены дремлющими программами, адаптированными к зеркальному письму. Попытайтесь написать какое‑нибудь слово в зеркальном отражении. Вероятно, вы потерпите неудачу. Теперь спрячьте лист бумаги под стол и попробуйте снова, как будто вы видите через столешницу. Вы увидите, что зеркальные жесты происходят совершенно естественно. Если вы снимете свою руку на видео и воспроизведете запись, то убедитесь, что она движется именно так, как нужно, чтобы писать справа налево. Аналогичным образом ваша правая рука может писать слева направо, а левая – справа налево. Одновременно. Писать зеркально обычно легче левой рукой (для правшей), что подтверждает гипотезу о положительной роли межполушарного переноса.
Наиболее убедительная поддержка дремлющей репрезентации зеркального чтения и письма наблюдается у больных с повреждениями мозга. Примерно у одного пациента с гемиплегией[463] из 40 имеются временные признаки зеркального письма. Этот любопытный симптом возникает почти всегда, когда болезнь вынуждает правшу писать левой рукой. В большинстве случаев дефицит касается только письма, но не чтения: обычно пациент не может прочитать зеркальные слова, которые он написал. Однако в настоящее время известно несколько человек, у которых после травмы мозга развилось одновременно и зеркальное письмо, и зеркальное чтение.

Рис. 7.7. Некоторые пациенты с повреждениями головного мозга внезапно начинают читать и писать в зеркальном отражении. На рисунке показаны образцы письма пациентки Х. Н. до и после автомобильной аварии (по материалам статьи Gottfried, Sancar, & Chatterjee, 2003). Используется с разрешения Neuropsychologia
Таков, например, случай с миссис Х. Н., 52‑летней американкой, попавшей в автомобильную катастрофу[464]. После аварии миссис Х. Н. страдала умеренной невнимательностью, а также нарушениями памяти и способности планировать, связанными с двусторонним снижением метаболизма лобных долей. Но главная ее жалоба заключалась в том, что она больше не могла ни читать, ни писать. Однажды, когда женщина ехала на машине, она заметила дорожный знак, на котором было написано «ПОТС». Оказалось, ей гораздо легче читать и писать в зеркальном отражении. Это удивительное наблюдение было подтверждено лабораторными экспериментами. Независимо от того, какой рукой пользовалась миссис Х. Н., она совершала гораздо больше ошибок, когда писала слева направо, чем справа налево (рис. 7.7).
Что еще удивительнее, обычные слова миссис Х. Н. читала хуже и медленнее, чем их зеркальные отражения. Распознавание слова, написанного справа налево, занимало у нее одну секунду, что в целом соответствует норме. Однако когда оно было написано правильно, слева направо, на эту же самую задачу женщине требовалось более двух с половиной секунд, что медленнее нормы в четыре раза. То же касалось слов, мелькавших на мониторе компьютера. При предъявлении зеркально перевернутой буквенной цепочки на четверть секунды миссис Х. Н. без труда определяла, английское это слово или нет. Как вы можете убедиться сами, это очень быстро.[465]

Как ни странно, когда слова были написаны правильно, слева направо, пациентка давала ответы наугад, причем даже в тех случаях, если длительность их предъявления составляла до половины секунды. Авария, в которую попала миссис Х. Н., повлекла за собой серьезные нарушения чтения и письма, а также обнажила дремлющую репрезентацию зеркальных отражений слов, о которой женщина даже не подозревала.
Случай миссис Х. Н. отнюдь не единичный. В неврологических журналах описано несколько таких пациентов. К сожалению, этого недостаточно, чтобы сделать окончательные выводы о лежащих в основе этого явления нейронных механизмах[466]. Повреждения мозга вследствие травмы головы часто размыты и плохо локализованы. Вероятно, для возникновения синдрома зеркального чтения необходима одновременная травматизация нескольких областей мозга. Было бы также преувеличением утверждать, чт
|
|
|

Адаптации растений и животных к жизни в горах: Большое значение для жизни организмов в горах имеют степень расчленения, крутизна и экспозиционные различия склонов...

Индивидуальные очистные сооружения: К классу индивидуальных очистных сооружений относят сооружения, пропускная способность которых...

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰)...
© cyberpedia.su 2017-2026 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!

