АТФ – является макроэргическим соединением, содержащим макроэргические связи; при гидролизе концевой фосфатной связи выделяется около 20 кдж/моль энергии.
К макроэргическим соединениям относятся ГТФ, ЦТФ, УТФ, креатинфосфат, карбамоилфосфат и др. Они используются в организме для синтеза АТФ. Например, ГТФ + АДФ à ГДФ + АТФ
Этот процесс называется субстратное фосфорилирование – экзоргонические реакции. В свою очередь все эти макроэргические соединения образуются при использовании свободной энергии концевой фосфатной группы АТФ. Наконец, энергия АТФ используется для совершения различных видов работ в организме:
- механической (мышечное сокращение);
- электрической (проведение нервного импульса);
- химической (синтез веществ);
-осмотической (активный транспорт веществ через мембрану) – эндергонические реакции.
Таким образом, АТФ- главный, непосредственно используемый донор энергии в организме. АТФ занимает центральное место между эндергоническими и экзергоническими реакциями.
В организме человека образуется количество АТФ, равное массе тела и за каждые 24 часа вся эта энергия разрушается. 1 молекула АТФ «живет» в клетке около минуты.
Использование АТФ как источника энергии возможно только при условии непрерывного синтеза АТФ из АДФ за счет энергии окисления органических соединений. Цикл АТФ-АДФ – основной механизм обмена энергии в биологических системах, а АТФ – универсальная «энергетическая валюта».
Каждая клетка обладает электрическим зарядом, который равен
[АТФ] + ½[АДФ]
[АТФ] + [АДФ] + [АМФ]
Если заряд клетки равен 0,8-0,9, то в клетке весь адениловый фонд представлен в виде АТФ (клетка насыщена энергией и процесс синтеза АТФ не происходит).
По мере использования энергии, АТФ превращается в АДФ, заряд клетки становится равным 0, автоматически начинается синтез АТФ.
Катаболизм – процесс расщепления органических веществ до конечных продуктов (СО2 , Н2О и мочевины). В этот процесс включаются метаболиты, образующиеся как при пищеварении, так и при распаде структурно-функциональных компонентов клеток.
Процессы катаболизма в клетках организма сопровождаются потреблением кислорода, который необходим для реакций окисления. В результате реакций катаболизма происходит выделение энергии (экзергонические реакции), которая необходима организму для его жизнедеятельности.
Анаболизм – синтез сложных веществ из простых. В анаболических процессах используется энергия, освобождающаяся при катаболизме (эндергонические реакции).
Цикл трикарбоновых кислот (ЦТК). Последовательность реакций. Связь с процессами окислительного фосфорилирования. Понятие о субстратном фосфорилировании. Энергетический эффект. Функции ЦТК.

Цикл Кребса открыт Гансом Кребсом в 1937г. (цикл трикарбоновых кислот), за открытие этого цикла Кребс в 1953г. получил Нобелевскую премию.
В ЦТК включается ацетил-КоА, который образуется в результате окисления жирных кислот, отдельных аминокислот и др. Некоторые метаболиты (глюкоза, глицерин и др.) превращаются в ПВК, из которой образуется ацетил-КоА в процессе окислительного декарбоксилирования:
пируватдегидрогеназный
комплекс
П  ВК ацетил-КоА
ВК ацетил-КоА
НАД (РР), ФАД (В2), ТДФ (В1),
HS- КоА (В3), липоевая кислота
Функции ЦТК.
Интегративная функция (объединяющая обмен белков, жиров и углеводов через образование ацетил-КоА).
Водородгенерирующая функция – образуется 4 пары протонов Н+.
Энергетическая функция – образуется 1 молекула АТФ между сукцинил-КоА и сукцинат (субстратное фосфорилирование).
Анаболическая (пластическая) – метаболиты ЦТК используются для синтеза других веществ:
- ацетил-КоА – в синтезе холестерина и его производных (желчных кислот, стероидных гормонов, провитамина Д3), высших жирных кислот, кетоновых тел, ацетилхолина и др;
- α-кетоглутаровая кислота – глутаминовая, глутамин, ГАМК, пролин, аргинин;
- сукцинил-КоА – гем;
- СО2 – глюкоза, высшие жирные кислоты, пуриновые и пиримидиновые нуклеотиды;
- фумаровая кислота – в синтезе мочевины;
- ЩУК – аспарагиновая кислота, аспарагин, глюкоза, метионин, треонин.
Субстраты ЦТК: изоцитрат, α-кетоглутарат и малат отдают протоны и электроны на НАД-зависимые дегидрогеназы, которые транспортируют протоны и электроны в ЦПЭ на I комплекс, следовательно протоны и электроны проходят все три пункта сопряжения (I, III, IV комплексы). Коэффициент окислительного фосфорилирования для:
-  изоцитрат – 3/1= 3АТФ;
изоцитрат – 3/1= 3АТФ;
- α-кетоглутарат – 3/1= 3АТФ; 9АТФ в ЦПЭ
- малат – 3/1= 3АТФ
Субстрат ЦТК: сукцинат отдает протоны и электроны на сукцинат-фумарат-дегидрогеназу (II комплекс), которая переносит протоны и электроны на убихинон, минуя I комплекс ЦПЭ, следовательно, электроны проходят два пункта сопряжения (III, IV комплексы).
Коэффициент окислительного фосфорилирования для сукцинат – 2/1= 2АТФ.
Между сукцинил-КоА и сукцинат образуется 1 молекула АТФ (субстратное фосфорилирование).
Энергетический баланс ЦТК= 9АТФ (ЦПЭ) + 2АТФ (ЦПЭ) + 1АТФ (субстратное фосфорилирование) = 12 молекул АТФ.
4.6. Биологическое окисление, особенности. Структурная организация дыхательной цепи. Последовательность расположения комплексов. Структура и роль их компонентов (ФМН, убихинон, цитохромы). Понятие о редокс-потенциалах.
Биологическое окисление – совокупность протекающих в каждой клетке ферментативных процессов, в результате которых молекулы углеводов, жиров и аминокислот расщепляются, в конечном счете, до углекислоты и воды, а освобождающаяся энергия запасается клеткой в виде аденозинтрифосфорной кислоты (АТФ) и затем используется в жизнедеятельности организма (биосинтез молекул, процесс деления клеток, сокращение мышц, активный транспорт, продукция тепла и др.).
Особенности биологического окисления:
4.
Протекает при температуре тела;
5.
В присутствии Н2О;
6.
Протекает постепенно через многочисленные стадии с участием ферментов-переносчиков, которые снижают энергию активации, происходит уменьшение свободной энергии, в результате чего энергия выделяется порциями. Поэтому окисление не сопровождается повышением температуры и не приводит к взрыву.
Электроны, поступающие в ЦПЭ, по мере их продвижения от одного переносчика к другому теряют свободную энергию. Значительная часть этой энергии запасается в АТФ, а часть рассеивается в виде тепла.
Перенос электронов от окисляемых субстратов к кислороду происходит в несколько этапов. В нем участвует большое количество промежуточных переносчиков, каждый из которых способен присоединять электроны от предыдущего переносчика и передавать следующему. Так возникает цепь окислительно-восстановительных реакций, в результате чего происходят восстановление О2 и синтез Н2О.
Убихинон - кофермент Q. Это небелковый переносчик. Он является жирорастворимым соединением. Структура КоQ сходна с витаминами К, Е. Убихинон выполняет коллекторную функцию, присоединяя ē и протоны от НАДН-дегидрогеназы (I комплекс), сукцинат-фумаратдегидрогеназы (II комплекс) и ФАД- зависимых дегидрогеназ, он обратимо восстанавливается в гидрохинон (QН2).Содержание убихинона значительно превосходит количество других компонентов ЦПЭ. Например, на 1 молекулу НАД+ приходится 50молекул КоQ.От убихинона происходит транспорт только электронов на цитохромы.
Цитохромы имеют ряд особенностей:
1)Цитохромы в ЦПЭ располагаются в порядке возрастания окислительно-восстановительного потенциала (редокс-потенциала);
2)Железо в цитохромах способно изменять свою степень окисления, поэтому цитохромы в ЦПЭ транспортируют только электроны.
В транспорте двух электронов принимают участие две молекулы каждого вида цитохромов, так как одна молекула цитохрома может переносить только один электрон.
5.6.Окислительное фосфорилирование. Сопряжение окисления (дыхания) и фосфорилирования (теория Митчелла).Vкомплекс-АТФ-аза. Коэффициент окислительного фосфорилирования Р/О. Дыхательный контроль. Так как электроны всегда стремятся переходить от электроотрицательных систем к электроположительным, их транспорт по ЦПЭ к кислороду сопровождается относительно большим снижением свободной энергии.
В ЦПЭ можно выделить 3 участка, в которых перенос электронов сопровождается относительно большим снижением свободной энергии. Это количество свободной энергии необходимо для синтеза АТФ из АДФ и фосфата (фосфорилирование).
Процесс переноса электронов по ЦПЭ и синтез АТФ энергетически сопряжены.
Синтез АТФ из АДФ и Н3РО4 за счет энергии переноса электронов по ЦПЭ называют окислительным фосфорилированием.
Механизм сопряжения окончательно не выяснен, наиболее обоснованной является хемиосмотическая теория Митчелла, предложенная в 1961г.
Перенос электронов по ЦПЭ от НАДН к кислороду сопровождается выкачиванием протонов из матрикса митохондрий через внутреннюю мембрану в межмембранное пространство.
Протоны, перенесенные из матрикса в межмембранное пространство, не могут вернуться обратно в матрикс, так как внутренняя мембрана не проницаема для протонов.
Таким образом, создается протонный градиент, при котором концентрация протонов в межмембранном пространстве больше, а рН меньше, чем в матриксе. Кроме того, каждый протон несет положительный заряд, и вследствие этого появляется разность потенциалов по обе стороны мембраны: отрицательный заряд – на внутренней стороне, положительный заряд – на внешней. В совокупности электрический и концентрационный градиенты составляют электрохимический потенциал ΔμН + - источник энергии для синтеза АТФ.
Энергия электрохимического потенциала (ΔμН+) используется для синтеза АТФ, если протоны возвращаются в матрикс через ионные каналы АТФ-синтазы (V комплекс).
Наиболее активный транспорт протонов в межмембранное пространство, необходимый для образования ΔμН+ происходит на участках ЦПЭ, соответствующих расположению комплексов I, III, IV. Эти участки называют пунктами сопряжения дыхания и фосфорилирования, где и происходит синтез АТФ.
V комплекс – АТФ-синтаза.
Это интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи. АТФ-синтаза состоит из двух белковых комплексов.
Гидрофобный комплекс F◦ погружен в мембрану. Он служит основанием, которое фиксирует АТФ-синтазу в мембране.
Комплекс F0 состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс.
Комплекс F1 выступает в матрикс. Он состоит из 9 субъединиц. Между α- и β- субъединицами располагаются три активных центра, в которых происходит синтез АТФ.
Повышение концентрации протонов в межмембранном пространстве активирует АТФ-синтазу. Электрохимический потенциал ΔμН+ заставляет протоны двигаться по каналу АТФ-синтазы в матрикс. Параллельно под действием ΔμН+ происходят конформационные изменения в парах α- и β- субъединиц белка F1 , в результате чего из АДФ и Н3РО4 образуется АТФ. Электрохимический потенциал, генерируемый в 3 пунктах сопряжения в ЦПЭ, используется для синтеза одной молекулы в каждом пункте.
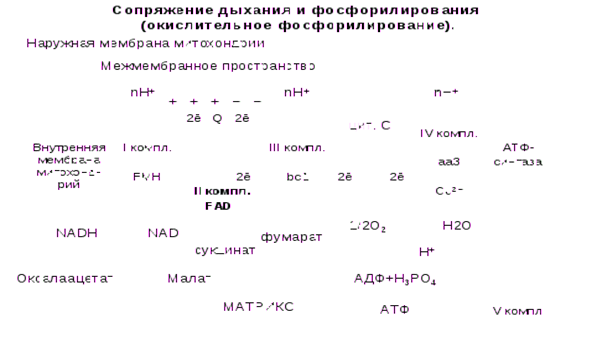
Отношение количества Н3РО4, использованной на фосфорилирование АДФ, к атому поглощенного кислорода, называют коэффициентом окислительного фосфорилирования и обозначают Р/О.
Для субстратов, которые отдают протоны и электроны на НАД-зависимые дегидрогеназы, коэффициент фосфорилирования равен 3/1=3АТФ, так как протоны и электроны транспортируются через 3 пункта сопряжения (I, III, IV комплексы).
Например, изоцитрат, малат, ПВК и др.
Для субстратов, которые отдают протоны и электроны на ФАД-зависимые дегидрогеназы и сукцинат-фумарат-дегидрогеназу (II комплекс), коэффициент фосфорилирования равен 2/1=2АТФ, так как электроны поступают на III комплекс, минуя первый пункт сопряжения (I комплекс). Например, сукцинат, глицерол-3-фосфат и др.
Зависимость интенсивности дыхания митохондрий от концентрации АДФ называют дыхательным контролем.
Разобщение окисления и фосфорилирования, разобщающие факторы. Биологическая роль термогенина бурой жировой ткани в детском организме. Гипоэнергетические состояния как результат гипоксии, голодания, авитаминозов и других. причин. Микросомальное окисление. Биологическая роль.
На синтез молекулы АТФ расходуется примерно 40-45% всей энергии электронов, переносимых по ЦПЭ, 25% тратится на работу по переносу веществ через мембрану. Остальная часть энергии рассеивается в виде теплоты и используется на поддержание температуры тела. Дополнительное образование теплоты происходит при разобщении дыхания и фосфорилирования, которое может быть биологически полезным. Оно позволяет генерировать тепло для поддержания температуры у новорожденных, у зимнеспящих животных и у всех млекопитающих в процессе адаптации к холоду. У них существует бурый жир – особая ткань, специализирующаяся на теплопродукции посредством разобщения дыхания и фосфорилирования. Бурый жир содержит много митохондрий. Около 10% всех белков приходится на так называемый разобщающий белок (РБ-1) – термогенин.
Окисление субстратов и фосфорилирование АДФ в митохондриях прочно сопряжены. Скорость использования АТФ регулирует скорость потока электронов в ЦПЭ. Если АТФ не используется и его концентрация в клетках возрастает, то прекращается и поток электронов к кислороду. С другой стороны, расход АТФ и превращение его в АДФ увеличивает окисление субстратов и поглощение кислорода.
Зависимость интенсивности дыхания митохондрий от концентрации АДФ называют дыхательным контролем.
Некоторые химические вещества (протонофоры) могут переносить протоны и другие ионы (ионофоры) из мембранного пространста через мембрану в матрикс, минуя протонные каналы АТФ-синтазы. В результате этого исчезает электрохимический потенциал и прекращается синтез АТФ. Это называют разобщением дыхания и фосфорилирования. В результате разобщения количество АТФ снижается, а АДФ увеличивается, что приводит к возрастанию скорости окисления НАДН и ФАДН2, возрастает и количество поглощенного кислорода, но энергия выделяется в виде тепла, и коэффициент Р/О резко снижается (свободное окисление).
Разобщители – липофильные вещества, легко проходящие через липидный слой мембраны:
- 2,4-динитрофенол,
- дикумарол (антивитамин вит. К);
- билирубин (продукт распада гема);
- тироксин (гормон щитовидной железы).
Все эти вещества проявляют разобщающее действие только при их высокой концентрации.
Для постоянного синтеза АТФ клетками необходим приток метаболитов как субстратов дыхания и кислорода как конечного акцептора электронов в реакциях окисления, сопряженных с синтезом АТФ.
Нарушения какого-либо этапа метаболизма, приводящие к прекращению синтеза АТФ, гибельны для клетки.
Состояния, при которых синтез АТФ снижен, объединяют термином «гипоэнергетические».
Причины «гипоэнергетических состояний»:
Голодание (недостаток субстратов дыхания);
Гиповитаминозы В1, В2, В3, РР – эти витамины входят в состав ферментов, участвующих в транспорте электронов на кислород.
Гипоксия может возникнуть: при недостатке кислорода во вдыхаемом воздухе; при заболеваниях легких и нарушении легочной вентиляции; при нарушениях кровообращения, вызванных заболеваниями сердца, спазмом и тромбозом сосудов, кровопотерей; при наследственных и приобретенных нарушениях структуры гемоглобина (анемии) и др.
Протекает в микросомах эндоплазматического ретикулума.
10% молекулярного кислорода не является акцептором протонов Н+ , а непосредственно включается в молекулу субстрата;
В процессе микросомального окисления АТФ не синтезируется.
Функции.
Детоксикационная (обезвреживающая) функция. В реакциях микросомального окисления субстраты гидроксилируются, повышается их растворимость, что способствует их выведению или снижению их токсичности (вывод лекарственных веществ из организма, избытка холестерина и т.д.);
Пластическая функция – синтез различных соединений:
- из холестерина образуются желчные кислоты, стероидные гормоны (гормоны коры надпочечников, половые гормоны);
- из пролина оксипролин;
- из лизина оксилизин;
- из фенилаланина тирозин и др.
В реакциях микросомального окисления участвуют ферменты- гидроксилазы (оксигеназы), которые делятся на монооксигеназы, катализирующие включение одного атома кислорода в молекулу субстрата и диоксигеназы- включение двух атомов кислорода в молекулу субстрата.
7.6.Образование токсических форм кислорода (супероксиданион, гидроксилрадикал, пероксид водорода), их повреждающее действие. Антиоксиданты (витамины Е,А,С, убихинон и др.) и антиоксидантные ферменты (супероксиддисмутаза, каталаза, пероксидаза).
В большинстве реакций с участием молекулярного кислорода его восстановление происходит поэтапно с переносом одного электрона на каждом этапе. При одноэлектронном переносе происходит образование промежуточных высокореактивных форм кислорода.
В невозбужденном состоянии кислород нетоксичен. Образование токсических форм кислорода связано с особенностями его молекулярной структуры. О2 содержит 2 неспаренных электрона с параллельными спинами, которые не могут образовывать термодинамическую стабильную пару и располагаются на разных орбиталях. Каждая из этих орбиталей может принять ещё один электрон.
Полное восстановление О2 происходит в результате 4 одноэлектронных переходов:
ē
О2 О2ˉ - супероксид
+ē, Н+
О2- Н2О2 - пероксид
+ē, Н+ +ē, Н+
Н2О2 Н2О + ОН- 2Н2О
гидроксильный радикал
Супероксид, пероксид и гидроксильный радикал- активные окислители, представляют серьезную опасность для многих структурных компонентов клетки.
Активные формы кислорода могут отщеплять электроны от многих соединений, превращая их в новые свободные радикалы, инициируя цепные окислительные реакции. Активные формы О2 вызывают окисление липидов, белков мембраны клеток, ДНК, РНК. Все это приводит к разрушению клеток.
Большая часть активных форм О2 образуется при переносе электронов в ЦПЭ, прежде всего, при функционировании QН2– дегидрогеназного комплекса. Это происходит в результате неферментативного переноса («утечки») электронов с QН2 на кислород.
Супероксид может образовываться:
1.
При спонтанном окислении гемоглобина. В норме гемоглобин обратимо связывает кислород в оксигемоглобин, однако оксигемоглобин превращается в метгемоглобин (Fе3+) и супероксид О2ˉ.
2.
Вирус гриппа взаимодействует с нейтрофилами легочной ткани и образует О2ˉ, который поддерживает воспаление, деструктивные процессы.
3.
Нейтрофилы, в избытке накапливающиеся в воспаленных суставах, также образуют О2ˉ, которые участвуют в развитии артритов.
В организме существуют защитные механизмы:
- гемсодержащие ферменты: пероксидаза, которая расщепляет Н2О2:
пероксидаза
Н  2О2 Н2О+О
2О2 Н2О+О
В организме существуют защитные механизмы:
- гемсодержащие ферменты: каталаза, которая расщепляет Н2О2:
каталаза
2  Н2О2 2Н2О + 2О2
Н2О2 2Н2О + 2О2
- супероксиддисмутаза (СД), глутатионпероксидаза, обезвреживающие О2ˉ:
СД
2  О2- + 2Н+ 2Н2О2 + О2
О2- + 2Н+ 2Н2О2 + О2
каталаза
2 Н2О2 2Н2О + 2О2
- антиоксиданты, обезвреживающие свободные радикалы – витамины Е, А, С; глутатион, цистеин, биофлавоноиды, Q10, мочевая кислота, янтарная кислота, селенит натрия и др.
1.7.Углеводы, классификация: моносахариды (глюкоза, фруктоза, галактоза, рибоза, и их производные (аминосахара, уроновые кислоты, фосфорные эфиры);дисахариды (сахароза, лактоза, мальтоза); гомополисахариды, гетерополисахариды. Химическое строение, биологическая роль.
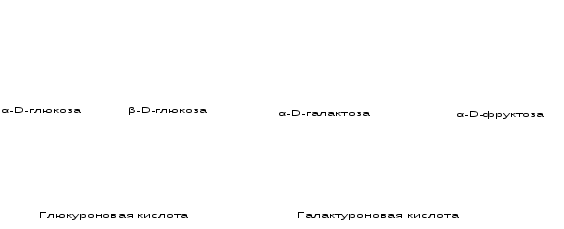
Углеводы вместе с белками, липидами и нуклеиновыми кислотами входят в состав живых организмов и определяют специфичность их строения и функционирования. На долю углеводов приходится около 75% массы пищевого рациона и более 50% от суточного количества необходимых калорий. Углеводы являются поставщиками энергии и выполняют структурную роль. Из углеводов в процессе метаболизма образуются вещества, которые служат исходными субстратами для синтеза липидов, аминокислот, нуклеотидов.
Углеводы – это альдегидо- или кетоноспирты.
Функции углеводов:
1.
Энергетическая;
2.
Углеводы (рибоза, дезоксирибоза) входят в состав нуклеиновых кислот, свободных мононуклеотидов (АТФ, ГТФ, цАМФ и др.), коферментов (НАД, НАДФ, ФАД);
3.
Структурная: гликопротеины – коллаген; белки-рецепторы; гликокаликс, белки определяющие принадлежность к группе крови; факторы свертывания крови; ферменты, гормоны; гликозаминогликаны и др;
4.
Защитная: иммуноглобулины, интерферон, муцины, фибриноген, гликозаминогликаны и др.;
5.
Дезинтоксикационная - входят в состав ФАФС (фосфоаденозинфосфосульфат) и УДФГК (уридиндифосфоглюкуроновая кислота).
Суточная потребность – 500 грамм.
Классификация:
1.
Моносахариды – производные многоатомных спиртов, содержащих альдегидную или кетонную группу. В
зависимости от количества углеродных атомов моносахариды делятся на:
Истинные:
- триозы
- тетрозы
- пентозы (рибоза, дезоксирибоза)
- гексозы (глюкоза, галактоза, фруктоза) и др.
Производные моносахаридов:
- уроновые кислоты – если вместо СН2ОН группы в 6 положении – СООН группа (у глюкозы – глюкуроновая
кислота; у галактозы – галактоуроновая кислота). Они входят в состав гликозаминогликанов;
- аровые кислоты – если в 6 и 1 положениях – СООН группы (у глюкозы – глюкаровая, у галактозы –
галактаровая);
- аминосахара – если во 2 положении – NН2группа (у глюкозы – глюкозамин, у галактозы – галактозамин). Они
входят в состав гликозамингликанов;
- N-гликозиды (рибоза, дезоксирибоза, соединенные с азотистым основанием в нуклеотиде - N-гликозидной
связью). Входят в состав РНК, ДНК, АТФ, НАД, ФАД и др.;
- фосфорные эфиры моносахаридов – замещение атомов Н на остатки фосфорной кислоты: глюкозо-6-фосфат;
фруктозо-1,6-дифосфат; рибозо-5-фосфат и др.
2. Олигосахариды (в их состав входят от 2 до 10 остатков моносахаридов)
Дисахариды:
- мальтоза состоит из 2 молекул α-Dглюкозы, соединенных α-1,4-гликозидной связью. Она имеет свободный
полуацетальный гидроксил, обладает восстанавливающими свойствами;
- лактоза (молочный сахар) состоит из β-Dгалактозы и α-Dглюкозы соединенные β-1,4-гликозидной связью.
Имеет свободный полуацетальный гидроксил и обладает восстанавливающими свойствами.
- сахароза состоит из α-Dглюкозы и β-Dфруктозы, соединенные α-1,2-гликозидной связью. Свободного
полуацетального гидроксила не имеет, поэтому не обладает восстанавливающими свойствами.
3. Полисахариды:
- гомополисахариды (крахмал, клетчатка, гликоген);
- гетерополисахариды (гликопротеины, протеогликаны, гликолипиды).).
2.7. Особенности переваривания и всасывания углеводов у детей. Роль клетчатки. Нарушения переваривания углеводов. Мальабсорбция. Лактазная недостаточность удетей, причины, проявления. Общая схема источников и путей расходования глюкозы в организме.
Попадая в желудочно-кишечный тракт, углеводы под действием ферментов распадаются на моносахариды и всасываются эпителиальными клетками тощей и подвздошной кишок с помощью специальных механизмов транспорта через мембраны этих клеток (путем облегченной диффузии и активного транспорта).
В ротовой полости пища измельчается при пережевывании, смачиваясь при этом слюной, рН которой равна 6,8. Под влиянием α-амилазы слюны (эндоамилаза) происходит расщепление в крахмале α-1,4-гликозидных связей. Она не расщепляет α-1,6-гликозидные связи в крахмале, поэтому крахмал переваривается лишь частично с образованием крупных фрагментов – декстринов и небольшого количества мальтозы. α-амилаза не гидролизует гликозидные связи в дисахаридах.
В желудке действие амилазы слюны прекращается, т.к. рН желудочного сока равен 1,5-2,5. Однако, внутри
пищевого комка активность амилазы может некоторое время сохранятся, пока рН не изменится в кислую
сторону.
В 12-перстной кишке рН равна 7,5-8,0. Из поджелудочной железы в кишечник поступает панкреатическая α-амилаза. Этот фермент также является эндогликозидазой т.к. расщепляет α-1,4-гликозидные связи в крахмале и декстринах. Продукты переваривания: олигосахариды, содержащие 3-8 остатков глюкозы, мальтоза, изомальтоза – дисахарид, состоящий из 2 молекул α-Dглюкозы, соединенных α-1,6-гликозидной связью. Дальнейшее их переваривание происходит в нижних отделах тонкого кишечника под действием мальтазы, изомальтазы. Дисахариды пищи сахароза и лактоза также расщепляются в тонком кишечнике сахаразой и лактазной (полостное пищеварение).
Процесс переваривания заканчивается на поверхности эпителиальных клеток кишечника (мембранное,
пристеночное пищеварение). Эпителиальные клетки покрыты микроворсинками, над которыми располагается
волокнистая сеть - гликокаликс (гликопротеин). В нем располагаются ферменты гидролизующие мальтозу,
сахарозу, лактозу, которые не расщепилисьв полости кишечника.
Скорость всасывания моносахаридов различна, глюкоза и галактоза всасываются быстрее, чем другие
моносахариды. Транспорт моносахаридов в клетке слизистой оболочке кишечника может осуществляться
различными способами: путем облегченной диффузии и активного транспорта. При высокой концентрации
глюкозы в просвете кишечника она транспортируется в клетку путем облегченной диффузии. При низкой
концентрации – глюкоза всасывается путем активного транспорта.
Механизм активного транспорта.
Глюкоза и Nа+ соединяются с разными участками белка-переносчика. При этом Nа+ поступает в клетку по градиенту концентрации и одновременно транспортируется глюкоза против градиента концентрации. Чем больше градиент Nа+, тем больше поступления глюкозы в энтероциты. Если концентрация Nа+ уменьшается, транспорт глюкозы снижается. Свободная энергия, необходимая для активного транспорта образуется благодаря гидролизу АТФ, связанному с натриевым насосом, который «откачивает» из клетки Nа+ в обмен на К+. Глюкоза соединяется с другим белком-переносчиком путем облегченной диффузии всасывается в кровь.
Мальабсорбция – группа заболеваний, связанная с нарушением:
1.
переваривания углеводов в ЖКТ (дефект ферментов);
2.
нарушение всасывания продуктов распада моносахаридов.
Примером первой группы заболеваний является лактазная недостаточность (дисахаридазная).
У детей различают 2 формы:
- транзиторная (до года жизни), связанная с незрелостью фермента лактазы;
- генетическая – мутация гена, ответственного за синтез фермента лактазы.
У взрослых:
- дефект лактазы вследствие экспрессии гена лактазы возрастного характера, при этом непереносимость молока у лиц африканского и азиатского происхожде6ния. Средняя частота данной формы в странах Европы – 7-12%, в Китае 80%, в отдельных районах Африки – 97% (исторически сложившийся рацион питания);
- приобретенного характера- при кишечных заболеваниях (гастриты, колиты, энтериты). Как известно, активность лактазы ниже, чем других дисахаридаз, поэтому понижение её активности становится более заметным.
Проявление во всех случаях: осмотическая диарея, которую вызывают нерасщепленные дисахариды и невсосавшиеся моносахариды, поступающие в дистальные отделы кишечника, изменяют осмотическое давление, частично подвергаются ферментативному расщеплению микроорганизмами, с образованием кислот, газов, усиливается приток воды в кишечник, увеличивается объем кишечного содержимого, увеличивается перистальтика, появляются метеоризм и боли.
3.7.Катаболизм глюкозы. Аэробный распад – основной путь катаболизма глюкозы у человека. Последовательность реакций, распространение и физиологическое значение аэробного распада глюкозы. АЭРОБНЫЙ ГЛИКОЛИЗ – процесс окисления глюкозы до СО2 и Н2О, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции находятся в цитозоле клетки.
Выделяют 2 этапа:
1.
Подготовительный – глюкоза фосфорилируется и расщепляется на 2 молекулы фосфотриоз. Используются 2 молекулы АТФ.
2.
Этап, сопряженный с синтезом АТФ. Фосфотриозы превращяются в ПВК, которая далее окисляется до СО2 и Н2О (ЦТК). Таким образом, выход АТФ – 38 молекул.
Основное физиологическое значение – использование энергии, которая освобождается в этом процессе для синтеза АТФ. Метаболиты гликолиза используются для синтеза новых соединений (фр-6-фосфат и глицеральдегид-3-фосфат участвуют в образовании рибозо-5-фосфата – структурного компонента нуклеозидов; 3-фосфоглицерат включается в синтез аминокислот: серин, глицин, цистеин). Являются субстратами для НАД-зависимых дегидрогеназ дых. цепи (глицеральдегид -3-фосфат, пируват, изоцитрат, α-кетоглутарат, малат). В печени и жировой ткани ацетил-КоА, образующийся из ПВК, используется как субстрат при биосинтезе жирных кислот, холестерина
4.7.Анаэробный распад глюкозы (гликолиз),значение анаэробного распада глюкозы. Эффект Пастера.
АНАЭРОБНЫЙ ГЛИКОЛИЗ – процесс расщепления глюкозы с образованием конечного продукта лактата. Он протекает без использования кислорода и поэтому не зависит от работы митохондриальной дыхательной цепи. АТФ образуется за счет субстратного фосфорилирования (2 АТФ). Протекает в мышцах, в первые минуты мышечной работы, в эритроцитах (в которых отсутствуют митохондрии), а также в разных органах в условиях ограниченного снабжения их кислородом, в том числе в опухолевых клетках. Этот процесс служит показателем повышенной скорости деления клеток при недостаточной обеспеченности их системой кровеносных сосудов.
Сходство анаэробного и аэробного гликолиза заключается в том, что до стадии образования ПВК эти процессы протекают одинаково при участии тех же ферментов.
СН3 ЛДГ СН3
Глюкоза С=О 2 Н-С-ОН
СООН СООН
ПВК 2 НАДН2 2НАД лактат
Энергетический баланс анаэробного окисления глюкозы: образование 2 АТФ.
Основное физиологическое значение – использование энергии, которая освобождается в этом процессе для синтеза АТФ. Метаболиты гликолиза используются для синтеза новых соединений (фр-6-фосфат и глицеральдегид-3-фосфат участвуют в образовании рибозо-5-фосфата – структурного компонента нуклеозидов; 3-фосфоглицерат включается в синтез аминокислот: серин, глицин, цистеин).
ЭФФЕКТ ПАСТЕРА. Заключается в угнетении дыханием (О2) анаэробного гликолиза, т.е. происходит переключение с аэробного гликолиза на анаэробное окисление. Если ткани снабжены О2, то 2НАДН2, образовавшийся в процессе центральной реакции оксидоредукции, окислится в дыхательной цепи, поэтому ПВК не превращается в лактат, а в ацетил-КоА, который вовлекается в ЦТК.
Биосинтез глюкозы (глюконеогенезе) из аминокислот, глицерина и молочной кислоты. Значение глюконеогенеза для детского организма. Взаимосвязь гликолиза в мышцах и глюконеогенеза в печени (цикл Кори). Гормональная регуляция.
Это процесс синтеза глюкозы из веществ неуглеводной природы. Процесс протекает в печени и менее интенсивно в корковом веществе почек, а также в слизистой кишечника. Эти ткани могут обеспечивать синтез 80-100 г глюкозы в сутки.
Первичные субстраты – лактат, аминокислоты, глицерол. Их включение в глюконеогенез зависит от физиологического состояния организма:
лактат – постоянно;
глицерол – высвобождается при гидролизе жиров в период голодания или при длительной физической нагрузке;
аминокислоты – образуются в результате распада мышечных белков и включаются в глюконеогенез при длительном голодании или продолжительной мышечной работе.
Если гликолиз протекает в цитозоле, а часть реакций глюконеогенеза происходит в митохондриях. Процесс идет обратно анаэробному гликолизу за исключением 3-х необратимых реакций:
1.
Пируваткиназная – образование фосфоенолпирувата из ПВК происходит в ходе 2-х реакций, первая из кот. протекает в митохондриях. ПВК транспортируется в матрикс митохондрий и там карбоксилируется с образованием оксалацетата (ЩУК). Фермент-пируваткарбоксилаза, коферментом которым является биотин. Реакция протекает с использование АТФ. Затем оксалацетат (в цитозоле) превращается в фосфоенолпируват входе реакции, катализируемой фосфоенолпируваткарбоксилазой – ГТФ-зависимым ферментом (-СО2, ГТФ → ГДФ).
2.
Фосфофруктокиназная: ФР-1,6-дифосфат →Фр-6-фосфат под действием фермента фруктозо-1,6-бифосфатаза.
3.
Гексокиназная: Гл-6-фосфат→ Глюкоза под действием фермента Гл-6-фосфатазы.
ЗНАЧЕНИЕ – поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок.
РЕГУЛЯЦИЯ: инсулин – тормозит синтез ферментов; глюкокортикоиды – индуцируют синтез ферментов; СТГ – повышает активность ферментов глюконеогенеза.
ЦИКЛ КОРИ.
(взаимосвязь гликолиза в мышцах и глюконеогенеза в печени).
Схема:

ЗНАЧЕНИЕ:
1.
Регуляция постоянного уровня глюкозы в крови.
2.
Обеспечивает утилизацию лактата.
3.
Предотвращает накопление лактата (снижение рН - лактоацидоз).
4.
Экономичное использование углеводов организмом.
5.
Регуляция обмена углеводов осуществляется на уровне тканей – кровь, печень, мышцы.
6.7. Представление о пентозофосфатном пути превращений глюкозы. Окислительная стадия (до образования рибозо-5-фосфат). Распространение и физиологическое значение у детей. Регуляция.
Пентозофосфатный путь является альтернативным путем окисления глюкозы. Это путь окисления глюкозы путем укорочения углеродной цепочки на один углеродный атом. Пентозофосфатный путь не приводит к синтезу АТФ.
В пентозофосфатном пути различают два этапа:
Окислительный путь включает две реакции дегидрирования с участием кофермента НАДФ и реакцию декарбоксилирования. В результате образуется НАДФН2 и пентозы (рибозро-5-фосфат).
Неокислительный путь служит для синтеза пентоз. Реакции этого пути обратимы, поэтому из пентоз могут образовываться гексозы. Промежуточные продукты (фруктозо-6-фосфат, глицеро-альдегид-3-фосфат) могут включаться в пути аэробного и анаэробного окисления. Знать химизм окислительного этапа.

Таким образом, в результате пентозофосфатного пути окисления глюкозы образуются пентозы, необходимые для синтеза нуклеиновых кислот (ДНК, РНК), коферментов (НАД, НАДФ, ФМН, ФАД), а также восстановленная форма НАДФ, водороды которого необходимы для восстановительных синтезов (синтез высших жирных кислот, холес







