Гипоксия, лекарственные средства, токсины, ионы кальция и магния, простагландины, заболевания, например, болезнь Паркинсона, миастения - это причины, вызывающие вышеперечисленные нарушения синаптической передачи. Анестезиологи умеют управлять функцией нервно-мышечных синапсов, например, издавна известно, что кураре (тубокурарин) давно употребляется для мышечного обездвиживания. Известно, что тубокурарин является миорелаксантом и вводится больным во время проведения искусственной вентиляции легких с целью выключения собственного дыхания и расслабления скелетных мышц на время оперативного вмешательства.
Действие миорелаксантов основано на механизмах, которые уже изучались студентами II курса в разделе физиология возбудимых мембран. Так все релаксанты скелетных мышц делятся на деполяризующие и недеполяризующие.
К деполяризующим относятся листенон, дитилин и другие. Механизм их действия связан с тем, что химическая структура их похожа на молекулу ацетилхолина. Поэтому при взаимодействии с Н-рецепторами постсинаптической мембраны эти лекарства вызывают ПД мышечных волокон, но ацетилхолинэстераза (обычно разрушающая ацетилхолин в синаптической щели) на листенон и другие миорелаксанты этого ряда не действует. В результате высокая концентрация миорелаксантов приводит к стойкой деполяризации концевых пластинок мышечных волокон и скелетные мышцы расслабляются. Разрушает деполяризующие миорелаксанты холинэстераза плазмы.
К недеполяризующим миорелаксантам относятся тубокурарин, мивакуриум и другие.
Механизм действия недеполяризующих миорелаксантов основан на конкуренции их с молекулами ацетилхолина, за места на сайтах холинорецепторов на концевой пластинке. Таким образом, недеполяризующие миорелаксанты блокируют действие ацетилхолина и предотвращают деполяризацию (или активацию) мышечного сокращения. Недеполяризующие миорелаксанты действуют также на рецепторы пресинаптической мембраны, препятствуя вхождению кальция, что вызывает блокирование выброса ацетилхолина.
Мышцы различаются по чувствительности к миорелаксантам: наиболее чувствительны глазные мышцы, ответственные за открывание и движение глаз; за ними следуют жевательные, шейные, мышцы конечностей, межреберные и абдоминальные. Диафрагма является наименее чувствительной мышцей, поэтому в качестве ранних признаков восстановления нейромышечной передачи отмечают икание и попытки дыхания.
Существует ряд заболеваний, связанных с нарушением функции синаптической передачи. Миастения (Myasthenia gravis). Заболевание проявляется в слабости проксимальных отделов мышц конечностей, которая усиливается при физической нагрузке мышц век, появлением птоза. В основе нарушений лежит снижение числа постсинаптических холинорецепторов. Белки (ацетилхолинорецепторы натриевых каналов) разрушаются собственными антителами, число которых резко увеличивается в крови больных миастенией. Миастения по современным представлениям является аутоиммунным заболеванием. Число ацетилхолинорецепторов на постсинаптической мембране уменьшается, амплитуда постсинаптических потенциалов концевой пластинки снижается, ПД на мышечных волокнах не возникает, сокращение не происходит, что проявляется в слабости и утомляемости скелетных мышц.
Значок Основываясь на знаниях этапов передачи химических сигналов в синапсах (образование, выделение,"судьба" ацетилхолина в синаптической щели и взаимодействие его с холинорецепторами на постсинаптической мембране), предложите лекарственное средство, с помощью которого можно лечить миастению и обоснуйте ваше предложение:________________________________________________________ _____________________________________________________________________________________________________________________________________________________.
Значок Сигнальные химические вещества, например, никотин, карбахолин оказывают на постсинаптическую мембрану такое же действие как и ацетилхолин. Особенность влияния этих соединений заключается в том, что они не разрушаются ферментом ацетилхолинэстеразой в синаптической щели. Поэтому никотин и подобные ему лекарства длительное время активируют постсинаптическую мембрану мышечного волокна. Эти знания дают основания врачам выбрать контролируемую дозу, например, никотина и применять их, например, для восстановления двигательной активности парализованных мышц после перенесенного инсульта.
Другие химические вещества: эзерин, прозерин, неостигмин, физостигмин обладают антихолинэстеразным действием, они инактивируют ацетилхолинэстеразу, поэтому ацетилхолин в синаптической щели не гидролизуется, что резко увеличивает продолжительность действия ацетилхолина, выделяемого из нервных окончаний. Вследствие этого указанные лекарства активируют открытие натриевых каналов постсинаптической мембраны и инициируют сократительную деятельность мышцы. Из-за чего они в лечебных целях используются для стимуляции парализованных мышц.
Если же концентрация ацетилхолина неуправляемо возрастает под действием перечисленных лекарств стимуляция мышечных волокон резко возрастает, что может вызвать мышечный спазм.
Стрихнин блокирует тормозные синапсы, точнее угнетает действие тормозного медиатора глицина в спинном мозге. Поэтому активность возбуждающих медиаторов становится превалирующей, и это резко повышает возбудимость нейронов. Нейроны начинают генерировать частые ПД, что вызывает тонические мышечные сокращения.
Ботулотоксин является самым сильным органическим ядом, его вырабатывают бактерии Clostridum botulinum. Ботулотоксин препятствует высвобождению в синаптическую щель ацетилхолина из везикул, находящихся в пресинаптическом окончании. Токсин блокирует слияние везикул с пресинаптической мембраной и экзоцитоз пузырьков с ацетилхолином. Нервное волокно возбуждается, а передача химических сигналов на мышцу не происходит (ацетилхолин не выбрасывается в синаптическую щель). Ботулотоксин полностью выключает возникновение нормальных кальций-зависимых ПКП - мышца не сокращается. Зная механизм действия яда на нервно-мышечное соединение врачи могут управлять работой синапса с помощью ботулотоксина. Например, при усилении напряжения мышц на одной стороне шеи, туловища (мышечные дистонии) введение ботулотоксина приводит к расслаблению мышц по описанному выше механизму. В настоящее время ботулотоксин применяют и в косметологии для "разглаживания" морщин мимической мускулатуры. Опишите, почему"разглаживаются" мимические мышцы после введения ботулотоксина ___________________________________________ ______________________________________________________________________
____________________________________________________________________.
Серотонин как нейротрансмиттер в большом количестве высвобождается в синапсах нейронов восходящей ретикулярной формации (управляют активным бодрствованием) и лимбической системы (контролируют эмоциональное состояние). В норме при передаче ПД от одного нейрона другому серотонин выделяется в синаптическую щель и взаимодействует с рецепторами постсинаптической мембраны. Серотонин гидролизуется с помощью ферментов моноаминоксидазы (МАО) или катехол-О-метилтрансферазы (СОМТ) в синаптической щели. Кроме этого серотонин, удаляется из синаптической щели путем обратного захвата (реаптейк) через специальные рецепторы пресинаптической мембраны.
Курсивом выделены важные теоретические знания, которые служат основанием фармакологам и клиницистам управлять количеством серотонина в синаптической щели, то есть с помощью лекарственных средств увеличивать или уменьшать количество серотонина для связи его с рецепторами на постсинаптической мембране. Известно 16 типов серотонинергических рецепторов на постсинаптической мембране нейронов. После взаимодействия серотонина с сайтом на постсинаптическом рецепторе, открываются метаботропные, связанные с G -белком каналы или ионотропные (лигандзависимые Na+) каналы.
Некоторые психические заболевания сопровождаются уменьшением выделения серотонина в синаптическую щель, у таких больных снижается психическая и физическая активность, развивается депрессия, появляется чувство страха.
Значок! Коррекцию низкой активности серотонинергической системы можно провести 2- я путями, основываясь на вышеизложенных знаниях:
1. Блокировать с помощью лекарственных средств ____________________ системы в синаптической щели, напишите какие _________________________ и _____________________________.
2. Блокировать с помощью лекарственных веществ ____________________ захват серотонина пресинаптической _________________. Такие лекарства получили название антидепрессанты и широко применяются в психиатрической практике.
Рассмотрим еще один механизм интеграции на уровне одного нейрона, который получил название суммация.
2. Суммация (лат. summatio - сложение) - слияние локальных ответов на подпороговые стимулы. Помните! - при действии одиночного подпорогового стимула ПД не возникает. Различают два вида суммации: временную (последовательную) и пространственную (одновременную).
2.1. Временная суммация является результатом слияния постсинаптических локальных ответов, которые вызываются быстро следующими друг за другом несколькими последовательными афферентными стимулами (рис. 5). Обязательным условием данного вида суммации являются короткие интервалы между приходящими раздражениями. Стимулы должны приходить с таким интервалом, чтобы вызванные ими последующие локальные ответы могли суммироваться с предыдущими, которые не успели "затухнуть". Таким образом, временная суммация в синапсе позволяет отфильтровывать слабые сигналы, поступающие на нейрон. Разберем подробно механизм возникновения временной суммации. В ответ на одиночный афферентный стимул, идущий от нейрона к другому нейрону, в пресинаптической части синапса освобождается 1 квант нейромедиатора. При этом на постинаптической мембране нейрона, как вам уже известно из раздела 3, обычно возникает подпороговый потенциал (локальный ответ) величиной 0,1 – 0,2 мВ, недостаточной для генерации ПД. Чтобы величина локального ответа достигла критического уровня - порога возникновения ПД, она должна уменьшится примерно на 10 мВ (см. рис. 5). Для этого требуется суммация на посисинаптической мембране клетки многих подпороговых локальных ответов. Суммация является совокупным результатом действия на нейрон входных сенсорных стимулов. Суммация постинаптических потенциалов происходит на аксонном холмике нейрона, где возникает распространяющийся потенциал действия.
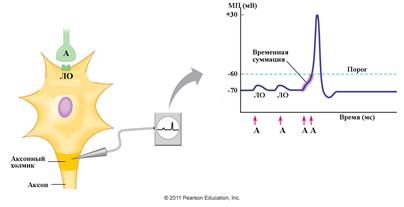
|
Рис. 5. Схема временной суммации. Обозначения: ЛО- локальный ответ (модифицировано по Pearson Education. Inc.)
2.2. Пространственная суммация связана со сложением локальных ответов, возникающих одновременно на разных участках одного нейрона (рис. 6). Механизм суммации аналогичен с рассмотренным в части 3. 1., с той лишь разницей, что на аксонном холмике генерируется ПД как результат слияния отдельных локальных ответов, возникающих одновременно на мембране одного нейрона, но на разных его элементах: теле, дендритах.
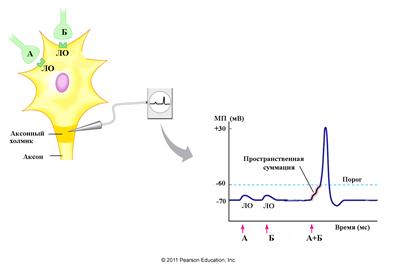
Рис. 6. Схема пространственной суммации. Обозначения: ЛО- локальный ответ (модифицировано по Pearson Education. Inc.)
Процессы интеграции в нервной системе происходят и при участие нескольких нейронов, рассмотрим несколько таких случаев:
1. Конвергенция, буквально означает схождение и предполагает передачу ПД от нескольких сенсорных нейронов к одному эффекторному (моторному) (рис.7, Б). Например, рефлекторное сгибание конечности в коленном суставе, можно вызвать путем раздражения различных рефлексогенных зон, поскольку на мотонейроне сходятся многие афферентные входы от различных рецепторов, располагающихся в коже, суставах, мышцах. Мотонейрон суммирует эти раздражения и отвечает двигательной реакцией.
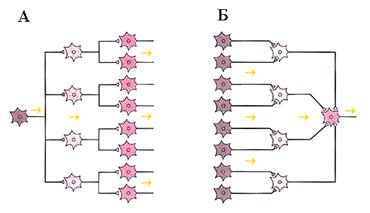
Рис. 7. Схема дивергенции - А и конвергенции - Б. Следует разделить эти рис на 2 и вставить в соответствующий текст и повернуть рис по часовой стрелке.
1.1. В связи с этим Чарльз Шеррингтон ввел понятие «общего конечного пути» или «принципа воронки». Общий конечный путь - принцип основанный на способности различных сенсорных путей создавать синаптические контакты на одной и той же эффекторной клетке. В тоже время принцип общего конечного пути показывает как раздражение различных структур мозга приводит к активации определенных групп мотонейронов. В описанном случае афферентные пути образуют подобие воронки, которая в узкой части сходится на мотонейроны (рис. 8).
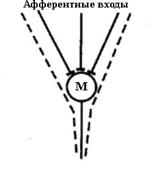
Рис.8. Схема, поясняющая принцип воронки Ч. Шеррингтона.
2. Дивергенция буквально означает расхождение (рис.7, А) ПД от одного сенсорного входа к другим нервным клеткам. Дивергенция вовлекает большое число нейронов в синхронную деятельность. Структурной основой дивергенции является значительное преобладание в нервной системе вставочных нейронов.
3. Иррадиация (лат. irradio - озарять, освещать) - широкое распространение процесса возбуждения по нейронам из одного участка нервной системе в другие. Иррадиация в нормально функционирующей нервной системе ограничивается тормозными процессами и другими. В незрелой нервной системе, в условиях гипоксии, наличия - возникновения патологических очагов раздражения возбуждение нерегулируемо рапространяется по нервной системе.
4. Окклюзия буквально обозначает "закупорка". Закупоривается (блокируется) возникновение ПД в нервном центре. Механизм окклюзии заключается в том, что при одновременном раздражении двух афферентных нервов 1 + 2 (рис. 9) суммарная активность нервных клеток, входящих в состав НЦ будет меньше, чем в случае раздражении 1 и 2 афферентных нервов порознь. Поясним сказанное на примере рисунка 9. При раздражении афферентного нерва 1 возбуждается 5 нейронов: А+Б+В+Г+Д, при раздражении афферентного нерва 2 возбуждается также 5 нейронов: Г+Д+Е+Ж+З. При раздражении 1 и 2 афферентных нервов одновременно активируются только 6 нейронов: А+Б+В+Е+Ж+З. Окклюзия происходит потому, что нейроны Г и Д являются общими для каждого из афферентных нервов.
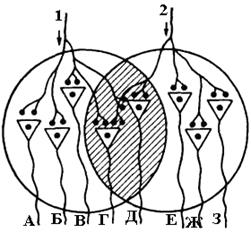
Рис. 9. Схема окклюзии в НЦ.
5. Трансформация ритма ПД в НЦ связана с тем, что нейроны снижают и/или повышают частоту приходящих к ним сигналов. Например, афферентные возбуждения приходят на нервную клетку с частотой 17 Гц, а на аксонном холмике нейрона в ответ генерируется 40 имп/с. Другой пример: частота входных раздражений 50 имп/с, а на выходе нейрона регистрируется 10 имп/с.
6. Высокая утомляемость НЦ объясняется расходом в химических синапсах, входящих в состав НЦ, нейромедиаторов.
7. Тонус НЦ представляет собой некоторый умеренный фоновый уровень спонтанной генерации ПД нервными клетками, которые входят в состав НЦ в отсутствии раздражителей. Происхождение тонуса НЦ связано с притоком на нейроны НЦ постоянных системных электрических и химических сигналов, действия локальных химических веществ: СО2, NO, аутакоидов и др..
8. Реверберация является примером кругового движения ПД в замкнутой локальной сети нейронов (рис....), когда возбуждение (ПД) возвращается к "первому" нейрону кольцевой цепочки. Лоренто де Но первым описал этот механизм циркуляции возбуждения в нервной системе. Реверберация обеспечивает задержку, усиление и длительное поддержание однажды возникшей собственной активности в круговой цепи нейронов по принципу положительной обратной связи. Считается, что этот механизм лежит в основе формирования кратковременной памяти, эмоций.
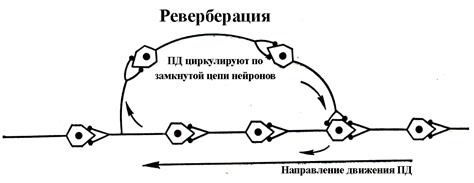
Рис.... Схема реверберации возбуждения. Дима, убери сверху рис надпись -Реверберация





