Строение синапса напоминает многоэтажный дом, состоящий из 2- х основных модулей (пре- и постсинаптическая мембраны) в которых конструкционным материалом являются специальные белки. Число их достигает 2000: структурный каркас пресинаптической мембраны насчитывает несколько сотен, а постсинаптической мембраны - около 1500. Прочный остов пре- и постсинаптических мембран образуют специальные цитоскелетные белки, адгезивные белки крепко связывают мембраны между собой. Другие белки выполняют функции ионных каналов, рецепторов, транспортеров и сигнальных молекул. Часть белков, входящих в молекулярную структуру мембран являются ферментными системами - киназы и фосфатазы. Важнейшими белками постсинаптической мембраны являются лиганд-управляемые и потенциал-зависимые каналы, а также транспортные белки.
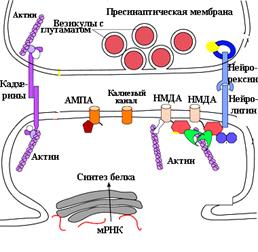
Рис.... Схема организации синапса в ЦНС. Обратите внимание, положение пре- и постсинаптической мембран между собой фиксировано с помощью белок-белковых соединений.
В состав центрального синапса входят 3-и обязательных (стандартных для всех синапсов) компонента.
Пресинаптическая мембрана включает синаптические везикулы, диаметром около 40 нм, митохондрии, белки адгезии и цитоскелета. В аксоплазме везикулы распределены неравномерно. Везикулы находятся в утолщенной части пресинаптической мембраны и хранят нейромедиатор. Везикулы сосредоточены в части, которая называется активная зона, в ней происходит слияние везикул с пресинаптической мембраной и высвобождение содержимого везикул (экзоцитоз) в синаптическую щель. Митохондрии поставляют АТФ для синтеза новых порций медиатора. Пресинаптическая мембрана включает значительное количество потенциалуправляемых кальциевых каналов.
Синаптическая щель (ширина которой составляет 20-25 нм) делит синапс на пре- и постсинаптические части.
Постсинаптическая мембрана является ограниченной частью постсинаптической нервной клетки, которая находится непосредственно в области контакта (через синаптическую щель) с активной зоной пресинаптической мембраны. Пре- и постсинаптические части мембран связаны между собой специальными адгезивными белками (Рис...). В постсинаптической мембране имеются специфические участки - большие белковые молекулы, которые насквозь пронизывают постсинаптическую мембрану (рис.. СДЕЛАТЬ). Структурно такой белок имеет несколько функциональных частей: одна часть (домен) трансмембранного белка выполняет роль рецептора, другая часть играет роль канала для движения разных ионов, существует еще один участок (домен) на этом белковом рецепторном комплексе, который находится внутри клетки (в цитоплазме), его активация приводит к метаболическим перестройкам в нейроне. Тот участок (домен) трансмембранного белка, который находится на внешней поверхности постсинаптической мембраны служит для связи с химическими молекулами - лигандами (нейротрансмиттерами из синаптической щели, гормонами, другими физиологически активными химическими молекулами). В центральном синапсе этот участок белкового рецептора играет роль замка, а молекула, например, нейромедиатора является ключом. Взаимодействие их (замка и ключа) "открывает" в белковой молекуле каналы для входа ионов в цитоплазму постсинаптического нейрона двумя путями. 1 - рецептор является частью каналообразующего белка, пронизывающего мембрану и канал открывается сразу же в результате связывания рецептора с лигандом. 2 - ионный канал открывается после активации системы вторичных посредников, находящихся внутри нейрона, последние управляют внутриклеточными ферментами, процессами трансдукции и пр.
Описанный выбор быстрого или медленного действия рецепторного белка на функцию нервных клеток зависит от типа рецепторных белков постсинаптической мембраны. Первый тип - ионотропные рецепторы постсинаптической мембраны. Они активируются нейромедиаторами. Стимуляция этих рецепторов вызывает открытие ионных каналов постсинаптической мембраны. Это быстродействующие рецепторы регулируют проницаемость ионного канала в течение миллисекунд после инактивации нейромедиатора из синаптической щели. Ионотропные рецепторы используются в ЦНС для передачи быстрых и адресных команд с одного нейрона на другой и с мотонейрона на скелетную мышцу.
Второй тип рецепторов именуется как метаботропные (ВЗЯТЬ РИС 45-7, со с. 623 Гайтона), и они очень чувствительны к воздействию многих химических молекул: нейромедиаторов, гормонов, физиологически активных веществ.Метаботропные рецепторы - медленнодействующие (от нескольких секунд до десятков дней), поэтому они участвуют, например, в регуляции функции внутренних органов, формировании памяти). Для будущего врача важно понимать, что теоретические знания о длительных по времени эффектах метаботропных рецепторов широко используются в практике фармакологами и фармацевтами для синтеза лекарственных средств пролонгированного действия, и применения таких фармакологических средств для эффективной терапии в клинике. Связывание метаботропного рецептора с лигандом (первичный посредник) вызывает образование вторичного посредника внутри цитоплазмы постсинаптического нейрона. Вторичные посредники изменяют обменные процессы в цитоплазме нервной клетки (отсюда и название рецепторов- метабо тропные).
Кратко остановимся на структуре и функциях вторичных посредников (мессенджеров). Выделяют несколько типов систем вторичных посредников: G- белки, цАМФ, цГМФ, ионы Са++, инозитол- 1,4,5- трифосфат (IP3) и диацилглицерин (DAG). Поясним на примере рисунка... внутриклеточную работу одного из вторичных посредников G- белка. К домену на внутриклеточной части мембраны прикреплен G- белок. В его состав входят три субъединицы: α- субъединица, β- и γ- субъединицы, последние две прикреплены как к α- субъединице, так и к внутренней стороне постсинаптической мембраны. Потенциал действия активирует α- субъединицу G- белка, которая отделяется от β- и γ- субъединиц. Альфа- субъединица свободно перемещается в цитозоле нейрона и реализовывает управляющие действия нейрона в зависимости от его функционального предназначения. На рис... показаны 4- е из вероятных функциональных влияний G- белка:
1 - Открытие специфических ионных каналов в постсинаптической мембране нейрона. Калиевый канал (позиция 1 рис....) открывается в ответ на воздействие α- субъединицы G- белка. Заметьте, К+- канал остается открытым в продолжении длительного времени, в отличие от быстрого закрытия ионного канала, в случае, когда G- белок (как вторичный посредник) не используется.
2- Активация циклического аденозинмонофосфата или циклического гуанозинмонофосфата в цитозоле нейрона (позиция 2 рис....). Взаимодействие нейромедиатора с рецептором на внешней стороне белковой молекулы активирует G- белок по схеме, описанной выше, что приводит в свою очередь к активации фермента аденилатциклазы. В результате катализа из АТФ образуется цАМФ (вторичный посредник), который активирует протеинкиназу А и инициируются многие эффекторные внутриклеточные реакции, включая длительные видоизменения структуры нейрона, что вызывает долговременные изменения возбудимости нервной клетки.
3- Активация одного или более из внутриклеточных ферментов. В этом случае G- белок по известной схеме активирует один или несколько внутриклеточных ферментов. Повышение активности ферментов приводит к усилению метаболических эффектов нейрона (позиция 3 рис....).
4- Активация генной транскрипции приводит к образованию новых эффекторных белков в нейроне и усиливает его структуру и функциональные возможности. При этом вторичные посредники активируются не только при участии G- белка, но и иными путями. Такие перестройки состава белков внутри нейрона определяют формирование долговременной памяти(позиция 4 рис....).
Важно! Независимо от типа рецептора постсинаптической мембраны, рецептор после связи с сигнальной химической молекулой побуждает нейрон к командному действию: активировать или тормозить постсинаптический нейрон, вызвать констрикцию или дилатацию гладкомышечных органов, секрецию гормонов и т.п.
После взаимодействия с рецептором излишки нейромедиатора удаляются во избегании продолжительного влияния на постсинаптическую мембрану. Иными словами "сделав свое дело" (помните, как мавр в трагедии У. Шекспира "Отелло") нейромедиатор должен "уйти". Известны 3- и механизма уменьшения концентрации нейромедиатора:
А- Элиминация (диффузия) из синаптической щели в окружающее нервные клетки пространство.
Б- Реаптейк (обратный захват) нейромедиатора пресинаптической мембраной.
3- Диссоциация нейромедиатора в синаптической щели с помощью ферментов.
В постсинаптических мембранах мозга имеется особая цитоскелетная структура - постсинаптическая плотность (ПСП) - сеть белковых филаментов и белковых модулей. Постсинаптическая плотность - это комплекс различных белков, структура которых динамически изменяется и зависит от активности данного синапса.
На электроннограмме ПСП определяется как электронно-плотный участок шириной 200-800 нм и толщиной 30-50 нм в непосредственной близи с постсинаптической мембраной (рис...). Ключевым формообразующим белком постсинаптической плотности является белок PSD - 95. В ПСП вcтроены (заякорены) ионотропные глутаматные рецепторы: AMPA и NMDA и калиевые каналы, а также кальций-кальмодулин-зависимая протеин киназа.. Эти рецепторные белки располагаются точно под пресинаптической мембраной, из которой везикулы выпускают глутамат и этот нейротрансмиттер путем диффузии достигает постсинаптической мембраны. Метаботропные (mGluRs) рецепторы находятся вне ПСП.
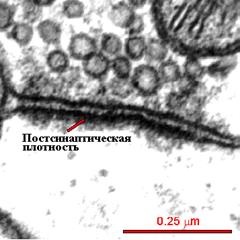
| Рис.... Электроннограмма постсинаптической плотности (по synapses.clm.utexas.edu с изменениями).
|
| | | | |
| | | | |
Указанные мембранные рецепторы и транспортеры, белки адгезии и цитоскелета необходимы для реализации нормальных функций синапса: распознавание и взаимодействие химического сигнала, активация канала и его конформация. Кроме этого ПСП регулирует размер и структуру дендритных шипиков. Ряд белков ППС, например, PDZ "заякоривают" мембранные рецепторы, "арматурные белки" GKAP, Shank и Homer обеспечивают встраивание разных подтипов рецепторов в определенные участки постсинаптической мембраны и регуляцию их количества и плотности. Иными словами, дизайн ПСП похож на многоэтажное здание, в котором на верхнем этаже располагаются постсинаптические рецепторы, ионные каналы, молекулы синаптической адгезии. На нижележащих этажах комплекс белков создает прочный каркас для физиологической регуляции числа рецепторов глутамата, которая зависит от функциональной активности данного синапса в передаче межнейронных химических сигналов. Таким образом ПСП обеспечивает кратковременную и длительную пластичность синапсов.
Линейные размеры ПСП сильно зависят от состояния активности синапсов. Изменение структуры постсинаптической плотности играет роль в формировании пластичности синаптической передачи (см. ниже).
С инапсы выполняют функцию усилителей сигналов при передаче ПД с постсинаптической мембраны на постсинаптическую.
В чем заключается суть усиления: один ПД инициирует выход из пресинаптической мембраны 100 везикул с 6000-8000 молекул нейротрансмиттера внутри каждой везикулы. Такой мощный химический сигнал воздействует на сравнительно небольшую по площади часть постсинаптической мембраны, на которой расположены рецепторы белковой природы (протеины), связывающиеся с молекулами нейротрансмиттера. Они вновь, но с усилением трансформируют химический сигнал в электрический на постсинаптической мембране. Усиление в химическом синапсе осуществляется также за счет участия вторичных посредников- активаторов метаболических процессов в клетках.
Описанные теоретические знания широко применяются в практической медицине. Например, применение многих современных лекарственных средств основано на знании механизмов управлении химическими сигналами в синапсах: усилении (облегчении условий передачи) или подавлении (вплоть до полной блокады).






