Клеточные мембраны. Химический состав. Пространственная организация и значение.
Клеточная мембрана, или плазмолемма – тонкая биологическая пленка, которая ограничивает клетку. Основу составляют двойной слой липидов. Липидный бислой содержит белки, которые делятся на 2 класса: 1) трансмембранные (их часть встроена в слой и пронизывает его насквозь); 2) периферические белки-рецепторы (расположены снаружи клеточной мембраны). Они покрыты слоем углеводов(образ. гликокаликс у животных клеток, а у растительных – целлюлозную оболочку.) Главная ф-ия – обеспечить поступление в клетку веществ и сохранить постоянство ее состава, то есть клеточный гомеостаз. Мембранный транспорт происходит путем эндоцитоза и экзоцитоза.
Химический состав, особенности морфологии хромосом. Динамика их структуры в клеточном цикле (Интерфазные и метафазные хромосомы).
Хромосомы – самовоспроизводящиеся структурные элементы клеточного ядра, обладающие особой организацией и функцией. Хромосома состоит из 2-ух плечей, которые разделены первичной перетяжкой - центромерой. От расположения центромеры различают 3 типа хромосом:
1. Метацентрические – первичная перетяжка находится в центре, плечи хромосомы равны.
2. Субметацентрические – первичная перетяжка сдвинута к краю, плечи неодинаковой длины.
3. Акроцентрические – первичная перетяжка расположена у самого края, одно плечо хромосомы очень короткое.
Концы плеч хромосом называются теломерами – это спец.участки, которые препятствуют соединению хромосом между собой. Каждая хромосома состоит из 2-ух хроматид. Хроматида состоит из двух спирально закрученных полухроматид. Поулхроматида состоит из двух скрученных фибрилл ДНК. По химическому составу хромосомы состоят из ДНК, белков, небольшого количества РНК, которые образуют нуклеопротеиновый комплекс. ДНК явл. носителем свойств наследственности и изменчивости. Хромосомные белки разделяются на: гистоны и негистоновые белки.
Гистоны представлены 5 фракциями: H1, H2a, H2в,H3, H4. Ф-ии: регуляторная(плотно связывается с ДНК и препятствует считыванию заключенной в ней биологической информации) и структурная(обеспечивает пространственную организацию ДНК в хромосомах).
Негистоновые белки состоят из ферментов синтеза и процессинга РНК, редупликации и репарации РНК. Ф-ии: структурная и регуляторная.
Интерфазные хромосомы деспирализованы (раскручены) и представляют собой не хромосомы (окрашенные тельца), а хроматин (окрашенные нити). В интерфазе происходит редупликация ДНК и транскрипция (синтез РНК), что возможно только тогда, когда хромосомы деспирализованы.
В профазе хроматин спирализуется, и в метафазе представляет собой уже хромосомы - окрашенные тельца определенной формы и размера, которые располагаются в экваториальной плоскости клетки, образуя метафазную пластинку.Спирализованное состояние облегчает прикрепление нитей ахроматинового веретена (= веретена деления) к центромерам (центральным перетяжкам хромосом) и расхождение хромосом в анафазе к полюсам клетки.
Генетический код, его свойства
Последовательность аминокислот в белке определяется последовательностью нуклеотидов в молекуле ДНК, его генетическим кодом. Т. о., наследственная информация на ДНК записана с помощью генетического кода.
Код имеет следующие основные свойства:
1. Триплетность — одну аминокислоту кодируют три нуклеотида.
2. Вырожденность (избыточность) — одну аминокислоту кодируют от двух до четырех триплетов. Всего имеется 64 триплета: 61 триплет несет информацию об аминокислотах, а 3 стоп-кодона обозначают окончание синтеза полипептидной цепи.
3. Неперекрываемость — нуклеотид одного триплета не может входить в состав соседнего триплета.
4. Универсальность — код един для всех живых организмов (синтез белка).
Размножение и его формы
Размножение – это всеобщее свойство живых организмов, заключающееся в способности производить подобных себе особей своего вида. Благодаря размножению происходит бесконечная смена поколений каждого вида. В процессе размножения могут возникать уникальные комбинации генетического материала, влекущие за собой появление наследственных изменений в организме. Таким образом, возникает генетическое разнообразие особей в пределах одного вида и закладываются основы изменчивости и дальнейшей эволюции вида.
Бесполое размножение. Древнейшей формой размножения на нашей планете является бесполое размножение. Оно заключается в делении одноклеточного организма (или одной или нескольких клеток многоклеточного организма) и образовании дочерних особей. Чаще эта форма размножения встречается у прокариот, растений, грибов и простейших, наблюдается она и у некоторых видов животных.
Виды бесполого размножения:
Размножение делением. У прокариот перед делением единственная кольцевая хромосома удваивается, между двумя дочерними хромосомами возникает перегородка и клетка делится надвое.
Многие одноклеточные водоросли (например, хламидомонада, эвглена зеленая) и простейшие (амеба) делятся митозом, образуя две клетки.
Размножение спорами. Споры – это специализированные гаплоидные клетки грибов и растений (не путать со спорами бактерий), служащие для размножения и расселения. У грибов и низших растений споры образуются путем митоза, у высших растений – в результате мейоза.
У семенных растений споры потеряли функцию расселения, но являются необходимым этапом цикла воспроизведения.
Вегетативное размножение. Представленные выше способы бесполого размножения объединяются тем, что новый организм во всех этих случаях развивается из одной клетки одноклеточного или многоклеточного родителя. Однако очень часто при бесполом размножении многоклеточных организмов потомство развивается из группы родительских клеток. Такой способ бесполого размножения называют вегетативным. Различают несколько видов вегетативного размножения. Первый из них – размножение растений частями вегетативных органов (часть слоевища, черенок стебля, черенок корня) или специальными видоизменениями побегов (корневище, луковица, клубень).
Другой вид вегетативного размножения – фрагментация, – процесс, основанный на регенерации. Так, например, фрагмент тела дождевого червя дает начало целой особи. Однако следует учитывать, что в природных условиях фрагментация встречается редко, в частности у многощетинковых червей, плесневых грибов, некоторых водорослей (спирогира).
Третий вид вегетативного размножения – почкование. В этом случае группа клеток родительской особи начинает согласованно делиться, давая начало дочерней особи, которая некоторое время развивается как часть материнского организма, а затем отделяется от него (пресноводная гидра) или формирует колонии из многих особей (коралловые полипы).
Значение бесполого размножения. Бесполое размножение позволяет быстро увеличивать численность особей данного вида в благоприятных условиях. Но при таком способе размножения все потомки имеют генотип, идентичный родительскому. Следовательно, при бесполом размножении практически не происходит увеличения генетического разнообразия, которое могло бы оказаться очень полезным при необходимости приспособиться к изменившимся условиям обитания. По этой причине подавляющее большинство живых организмов периодически или постоянно размножаются половым путем.
Встречается способ бесполого размножения, называемый шизогонией. Шизогония свойственна малярийным плазмодиям, относящимся к простейшим. При этом содержимое материнской клетки многократно делится и образуется множество новых клеток-паразитов.
Особым видом вегетативного размножения организмов является полиэмбриония. В этом случае зародыш (эмбрион) высших животных вскоре после образования делится на несколько фрагментов, каждый из которых независимо развивается в полноценную особь. Такое деление эмбрионов встречается, например, у броненосцев. К полиэмбрионии также относится образование однояйцевых близнецов у человека.
В этом случае зигота, возникшая в результате обычного оплодотворения, дробясь, образует зародыш, который, по пока не вполне понятным причинам, разделяется на несколько частей. Каждая из этих частей проходит путь нормального эмбрионального развития, в результате чего рождаются два и более практически одинаковых младенца, обязательно одного пола. Частота рождаемости однояйцевых близнецов не превышает одного случая на 250 обычных родов. Но иногда разделение формирующегося зародыша бывает неполным. В этом случае возникают организмы, имеющие общие части тела или внутренние органы.
15. Этапы гаметогенеза. Мейоз.
Гаметогенез – процесс развития половых клеток.
Стадии гаметогенеза
Размножение
Будущие половые клетки образуются из «заготовок» — специальных клеток с двойным (диплойдным) набором хромосом, которые называются овогонии(женские) и сперматогонии (мужские клетки).
И сначала эти клетки энергично делятся, делятся митозом, чтобы увеличить свой количество.
Интересно, что в мужском и женском организме этот период происходит в разное время.
Овогонии размножаются тогда, когда человека и женщиной-то назвать еще нельзя, это еще эмбрион. Т.е. женский организм рождается уже с определенным количеством овогоний. По истечении 7 месяцев развития эмбриона клетки приступают к мейозу. Сперматогонии размножаются в течение всего репродуктивного периода мужского организма. У всех организмов этот период разный, но, безусловно, он значительно дольше, чем у женского, и конечно, половых клеток в мужском организме образуется гораздо больше.
Рост
Рост, увеличение в размерах, гакопление питательных веществ — все это характеристики этапа роста, подготовки к делению — к мейозу 1. Именно на этой стадии эти клетки уже называют овоцитами и сперматоцитами I порядка.
Важно: на этом этапе кол-во хромосом остаеься то же, а вот ДНК удваивается!
Созревание
Собственно, мейоз — 2 последовательных деления: мейоз 1 и мейоз 2. И вот здесь имеет смысл разобрать отдельно образование женской половой клетки и мужской, потому что пути у них Очень разные.
Женская половая клетка:
· происходит мейоз 1 — количество хромосом уменьшается вдвое. Образуетсяовоцит II порядка.
· Второе деление — мейоз 2 — образуются четыре гаплойдные клетки: зрелую яйцеклетку и 3 редукционных тельца. В яйцеклетке будет больший запас питательных веществ. Она даст начало (при оплодотворении) новому живому организму. Поэтому такое неравномерное распределение обеспечивает поялание крупной, богатой питательными веществами клетки — яйцеклетки.На этом этапе гаметогенез для яйцеклетки закончен, и она готова выполнять свои функции — быть оплодотворенной мужской половой клеткой.

Мужская половая клетка:
· происходит мейоз 1 — количество хромосом уменьшается вдвое. Образуетсясперматоцит II порядка.
· Второе деление — мейоз 2 — образуются четыре гаплойдные клетки —сперматиды. Они переходят на 4-ю стадию процесса.
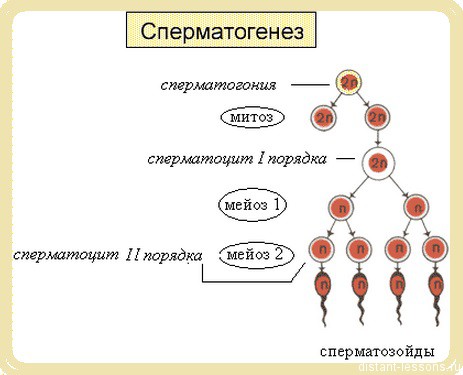
4. Формирование (спермиогенез)
Клетки «доформировываются». Им предстоит долгий и трудный путь до яйцеклетки. Победителем в этом марафоне будет только один, поэтому необходимо подготовиться: уплотняется ядро, хромосомы спирализуются, цитоплазма уходит; формируется жгутик — именно за счет него сперматозойдосущствляет поступательное движение, в нем должно быть много белки и митохондрии. Спринтер готов.
1. При гаметогенезе образуются половые клетки: мужские (сперматозойды) и женские (яйцеклетки).
2. И те, и другие содержат гаплойдный (1n) надор хромосом.
3. В результате гаметогенеза образуется 1 функциональная яйцеклетка и несколько сперматозойдов.
4. Овогенез и гаметогенез различаются по времени прохождения процессов и по количеству стадий.
Мейоз (от греч. мейозис — уменьшение) — это способ деления эукариотических клеток, в результате которого из одной материнской клетки образуются четыре дочерние с уменьшенным в 2 раза набором хромосом. Если в мейоз вступает диплоидная соматическая клетка (2п4с), то образуются четыре гаплоидные клетки (lnlc). Клетки с гаплоидным набором хромосом не могут делиться мейозом.
Мейоз представляет собой непрерывный процесс, состоящий из двух последовательных делений, называемых мейозом I и мейозом II. В каждом делении различают профазу, метафазу, анафазу и телофазу (рис. 58). В результате первого деления мейоза число хромосом уменьшается вдвое, в ходе второго деления мейоза гаплоидность клеток сохраняется.
В профазе мейоза I (или профазе I) растворяются ядрышки, распадается ядерная оболочка и начинается формирование веретена деления. Хроматин спи-рализуется с образованием двухроматидных хромосом (в диплоидной клетке — набор 2п4с). Гомологичные хромосомы попарно сближаются, этот процесс называется конъюгацией хромосом. При конъюгации хроматиды гомологичных хромосом в некоторых местах перекрещиваются. Между некоторыми хроматида-ми гомологичных хромосом может происходить обмен соответствующими участками — кроссинговер.
В метафазе I пары гомологичных хромосом располагаются в экваториальной плоскости клетки. В этот момент спирализация хромосом достигает максимума.
В анафазе I гомологичные хромосомы (а не сестринские хроматиды, как при митозе) отходят друг от друга и растягиваются нитями веретена деления к противоположным полюсам клетки. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадет только одна. Таким образом, в конце анафазы I набор хромосом и хроматид у каждого полюса делящейся клетки составляет \ti2c — он уже уменьшился вдвое, но хромосомы все еще остаются двухроматидными.
В телофазе I веретено деления разрушается, происходит формирование двух ядер и деление цитоплазмы. Образуются две дочерние клетки, содержащие гаплоидный набор хромосом, каждая хромосома состоит из двух хроматид (\п2с).
Промежуток между мейозом I и мейозом II очень короткий. И н т е р ф а з а II практически отсутствует. В это время не происходит репликация ДНК и две дочерние клетки быстро вступают во второе деление мейоза, протекающее по типу митоза.
В профазе II происходят те же процессы, что и в профазе митоза: формируются хромосомы, они беспорядочно располагаются в цитоплазме клетки. Начинает формироваться веретено деления.
В метафазе II хромосомы располагаются в экваториальной плоскости.
В анафазе II сестринские хроматиды каждой хромосомы разделяются и отходят к противоположным полюсам клетки. В конце анафазы II набор хромосом и хроматид у каждого полюса — \ti\c.
В телофазе II образуются четыре гаплоидные клетки, каждая хромосома состоит из одной хроматиды (lnlc).
Таким образом, мейоз представляет собой два последовательных деления ядра и цитоплазмы, перед которыми репликация происходит только один раз. Энергия и вещества, необходимые для обоих делений мейоза, накапливаются во время и н тер фазы I.
В профазе мейоза I происходит кроссинговер, что ведет к перекомбинации наследственного материала. В анафазе I гомологичные хромосомы случайным образом расходятся к разным полюсам клетки, в анафазе II то же самое происходит с сестринскими хроматидами. Все эти процессы обусловливают комби-нативную изменчивость живых организмов, о которой будет говориться позже.
Биологическое значение мейоза. У животных и человека мейоз приводит к образованию гаплоидных половых клеток — гамет. В ходе последующего процесса оплодотворения (слияния гамет) организм нового поколения получает диплоидный набор хромосом, а значит, сохраняет присущий данному виду организмов кариотип. Следовательно, мейоз препятствует увеличению числа хромосом при половом размножении. Без такого механизма деления хромосомные наборы удваивались бы с каждым следующим поколением.
У растений, грибов и некоторых протистов путем мейоза образуются споры. Процессы, протекающие в ходе мейоза, служат основой комбинативной изменчивости организмов.
Период старения человека.
Период старения у мужчин начинается после 60 лет, а у женщин после 55. По современной классификации людей, достигших 60 - 76 лет называют пожилыми, 75 - 89 старыми, а свыше 90 лет - долгожителями.
Старение представляет собой закономерную стадию индивидуального развития, свойственную всем живым организмам.
Регенерация и ее виды.
Регенера́ция — способность живых организмов со временем восстанавливать повреждённые ткани, а иногда и целые потерянные органы. Регенерацией также называется восстановление целого организма из его искусственно отделённого фрагмента (например, восстановление гидры из небольшого фрагмента тела или диссоциированных клеток).
Типы регенерации:
• Репаративной - называют регенерацию, происходящую после повреждения или утраты какой-либо части тела. Выделяют типичную и атипичную репаративнуюрегенерацию.При типичной регенерации утраченная часть замещается путём развития точно такой же части. Причиной утраты может быть внешнее воздействие (например, ампутация), или же животное намеренно отрывает часть своего тела (автотомия), как ящерица, обламывающая часть своего хвоста, спасаясь от врага.При атипичной регенерации утраченная часть замещается структурой, отличающейся от первоначальной количественно или качественно. У регенерировавшей конечности головастика число пальцев может оказаться меньше исходного, а у креветки вместо ампутированного глаза может вырасти антенна.
• Регенерацию в процессе нормальной жизнедеятельности организма, обычно не связанную с повреждениями или утратой, называют физиологической. В каждом организме на протяжении всей его жизни постоянно идут процессы восстановления и обновления. У человека, например, постоянно обновляется наружный слой кожи. Птицы периодически сбрасывают перья и отращивают новые, а млекопитающие сменяют шерстный покров. У листопадных деревьев листья ежегодно опадают и заменяются свежими. Такие процессы носят название физиологической регенерации.
•Внутриклеточная регенерация – процесс восстановления макромолекул и органелл. Увелечкние числа органелл достигается усилением их образования (лизосомы), сборки элементарных структурных единиц (микротрубочки), или путем их деления (митохондрии).
•Регенерация на тканевом, органном и организменном уровнях обеспечивается делением, детерминацией, дифференцировкой и ростом клеток.
Трансплантация — в медицине пересадка какого-либо органа или ткани, например, почки, сердца, печени, лёгкого, костного мозга, стволовых гемопоэтических клеток, волос.
Различают следующие виды трансплантации:
1.Аутотрансплантация, или аутологичная трансплантация — реципиент трансплантата является его донором для самого себя. Например, аутотрансплантация кожи с неповреждённых участков на обожжённые широко применяется при тяжёлых ожогах. Аутотрансплантация костного мозга или гемопоэтических стволовых клеток после высокодозной противоопухолевой химиотерапии широко применяется при лейкозах, лимфомах и химиочувствительных злокачественных опухолях.
2.Изогенная трансплантация — донором трансплантата является 100% генетически и иммунологически идентичный реципиенту однояйцевый близнец реципиента.
3.Аллотрансплантация, или гомотрансплантация — донором трансплантата является генетически и иммунологически отличающийся человеческий организм.
4.Ксенотрансплантация, или межвидовая трансплантация — трансплантация органов от животного другого биологического вида.
Основные понятия генетики
Наследственность — свойство организмов сохранять генетическую информацию, детерминирующую признаки, особенности развития и жизнедеятельность потомков. Наследственность закрепляет признаки, характерные для вида.
Наследование — способ передачи наследственной информации от одного поколения к другому.
Изменчивость — свойство организмов изменять признаки, полу-ченные от родителей, или приобретать новые в процессе индивиду-ального развития.
Ген — единица наследственности. В структурном отношении — это определенная последовательность нуклеотидов ДНК, кодирующая структуру полипептидной цепи. В функциональном отношении (как единица функции — цистрон) — это совокупность кодирующих и регуляторных последовательностей, одновременно участвующих в процессе транскрипции.
Гены располагаются в хромосомах линейно. Участок хромосомы, в котором расположен определенный ген, называется локусом. Гены одной хромосомы образуют группу сцепления и наследуются вместе. При кроссинговере сцепление генов нарушается.
Аллель — одно из возможных состояний гена (конкретное состо-яние кодирующей последовательности: определенный состав, по-следовательность и количество нуклеотидов в локусе).
Аллельные гены — гены, определяющие развитие одного и того же признака (цвет) в одинаковых или разных фенотипических про-явлениях (желтый, зеленый); локализованы в одинаковых локусах гомологичных хромосом.
Неаллельные гены — гены, определяющие развитие разных при-знаков (цвет и форма). Могут быть локализованы в гомологичных хромосомах, но в разных локусах, или в негомологичных хромосомах.
Гомозигота — организм, в гомологичных хромосомах которого находятся одинаковые аллели одного гена (изоаллели) — аа, АА, ААВВ, ааВВ; образует один тип гамет.
Гетерозигота — организм, в гомологичных хромосомах которого локализованы разные аллели одного гена (Аа, АаВв); образует разные типы гамет.
Гемизигота — организм, содержащий только один аллель определенного гена (мужской организм гемизиготен по генам, локализованным в X- и Y-хромосомах). Гены, расположенные в участке Y-хромосомы, не гомологичном Х-хромосоме, называются сцепленными с Y-хромосомой, а наследование — голандрическим.
Гибридизация — скрещивание гомозиготных особей, отличающихся по генотипу (фенотипу).
Рецессивный аллель — аллель, проявляющийся только у гомозиготных организмов (аа); у гетерозиготных подавляется доминантным аллелем.
Доминантный аллель — аллель, проявляющийся у гомо- и у ге-терозиготных организмов (АА, Аа).
Фенотип — совокупность всех признаков, свойств, особенностей развития и жизнедеятельности организма. Это результат реализации генотипа в конкретных условиях среды.
Генотип — система всех взаимодействующих аллелей организма.
Геном — количество генов в гаплоидном наборе, характерное для вида.
Моногибридное скрещивание — скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков. При этом скрещиваемые предки являются гетерозиготными по положению аллеля в хромосоме
Моногибридное наследование представляет собой пример наследования единственного признака (гена), различные формы которого называют аллелями. Например, при моногибридном скрещивании между двумя чистыми линиями растений, гомозиготных по соответствующим признакам -- одного с жёлтыми семенами (доминантный признак), а другого с зелёными семенами (рецессивный признак), можно ожидать, что первое поколение будет только с жёлтыми семенами, потому что аллель жёлтых семян доминирует над аллелью зелёных. При моногибридном скрещивании сравнивают только один характерный признак.
Законы Менделя — это принципы передачи наследственных признаков от родительских организмов к их потомкам, вытекающие из экспериментов Грегора Менделя. Эти принципы послужили основой для классической генетики и впоследствии были объяснены как следствие молекулярных механизмов наследственности. Хотя в русскоязычных учебниках обычно описывают три закона, «первый закон» не был открыт Менделем. Особое значение из открытых Менделем закономерностей имеет «гипотеза чистоты гамет».
Закон единообразия гибридов первого поколения, или первый закон Менделя, утверждает, что потомство первого поколения от скрещивания устойчивых форм, различающихся по одному признаку, имеет одинаковый фенотип по этому признаку. При этом все гибриды могут иметь фенотип одного из родителей (полное доминирование), как это имело место в опытах Менделя, или, как было обнаружено позднее, промежуточный фенотип (неполное доминирование). В дальнейшем выяснилось, что гибриды первого поколения могут проявить признаки обоих родителей (кодоминировапие). Этот закон основан на том, что при скрещивании двух гомозиготных по разным аллелям форм (АА и аа) все их потомки одинаковы по генотипу (гетерозиготны — Аа), а значит, и по фенотипу.
Закон расщепления, или второй закон Менделя, гласит, что при скрещивании гибридов первого поколения между собой среди гибридов второго поколения в определ. соотношениях появляются особи с фенотипами исходных родительских форм и гибридов первого поколения. Так, в случае полного доминирования выявляются 75% особей с доминантным и 25% с рецессивным признаком, т. е. два фенотипа в отношении 3:1 (рис. 1). При неполном доминировании и кодомииировании 50% гибридов второго поколения имеют фенотип гибридов первого поколения и по 25% — фенотипы исходных родительских форм, т. е. наблюдают расщепление 1:2:1. В основе второго закона лежит закономерное поведение пары гомологичных хромосом (с аллелями А и а), к-рое обеспечивает образование у гибридов первого поколения гамет двух типов, в результате чего среди гибридов второго поколения выявляются особи трёх возможных генотипов в соотношении 1АА:2Аа:1аа. Конкретные типы взаимодействия аллелей и дают расшепления по фенотипу в соответствии со вторым законом Менделя.
Признаки, которые наследуются по законам Менделя, называются менделирующими. Все менделирующие признаки дискретны и контролируются моногенно, то есть одним геном. Например, у человека к таким признакам относятся цвет глаз, острота зрения, наличие абсолютного слуха.
Ген, имеющийся в генотипе в необходимом для проявления количестве (1 аллель для доминантных признаков и 2 аллеля для рецессивных) может проявляться в виде признака в разной степени у разных организмов (экспрессивность) или вообще не проявляться (пенетрантность). Причины:
· модификационная изменчивость (воздействие условий окружающей среды)
· комбинативная изменчивость (воздействие других генов генотипа).
Экспрессивность – степень фенотипического проявления аллеля. Например, аллели групп крови АВ0 у человека имеют постоянную экспрессивность (всегда проявляются на 100%), а аллели, определяющие окраску глаз, – изменчивую экспрессивность. Рецессивная мутация, уменьшающая число фасеток глаза у дрозофилы, у разных особей по разному уменьшает число фасеток вплоть до полного их отсутствия.
Пенетрантность – вероятность фенотипического проявления признака при наличии соответствующего гена. Например, пенетрантность врожденного вывиха бедра у человека составляет 25%, т.е. болезнью страдает только 1/4 рецессивных гомозигот. Медико-генетическое значение пенетрантности: здоровый человек, у которого один из родителей страдает заболеванием с неполной пенетрантностью, может иметь непроявляющийся мутантный ген и передать его детям.
Правила наследования.
Правило чистоты гамет: Гены из поколения в поколение передаются в чистом виде, не смешиваясь и не изменяя, как правило, своей структуры.
Правило расщепления: При ди- и полигибридном скрещивании гены различных пар признаков комбинируются во всех возможных сочетаниях и во втором поколении дают расщепление по генотипу в соответствии с формулой (1:2:1)n, а по фенотипу (3:1)n,где n – число анализируемых признаков.
Система резус
Система резус контролируется тремя тесно сцепленными генами CDE, однако ген D является наиболее частой причиной иммунологического конфликта между матерью и плодам. Резус-конфликт между матерью и плодам возникает у 1 из 30 женщин в ситуации, когда мать резус-отрицательная, а плод резус-положительный.
Генотипы людей с положительным и отрицательным резусом
Rh+ Rh+, Rh+ Rh– - резус положительный
Rh– Rh– - резус отрицательный.
Во время беременности после 7-й недели, когда в крови плода появляются зрелые эритроциты, в организме матери начинают вырабатываться противорезусные антитела, которые, проникая через плаценту в кровяное русло плода, вызывает слипание (агглютинацию) эритроцитов и их разрушение.
Как правило, первая беременность заканчивается благополучно, т.к. противорезусные антитела не успевают накопиться в организме матери в достаточном количестве. Если после первых родов не проводилась соответствующая профилактика (введение сыворотки – анти-D-глобулина, связывающего резус-антиген), то при повторных беременностях повышается риск рождения ребенка с гемолитической болезнью новорожденных, проявляющейся анемией, желтухой, отеками и обуславливающей сложный дефект интеллекта, слуха и речи, двигательные расстройства.
25. Основные положения хромосомной теории наследственности.
Сцепление генов. Во всех примерах скрещивания, рассмотренных в предыдущих параграфах, имело место независимое комбинирование, поскольку интересующие нас пары генов находились в разных парах хромосом. Такие гены свободно сочетаются друг с другом в соответствии с закономерностью независимого комбинирования.
У различных организмов число генов может насчитывать десятки и сотни тысяч, число же хромосом весьма ограниченно, другими словами, число генов значительно превосходит число хромосом. Вот почему в каждой хромосоме располагается множество генов. Гены, локализованные в одной хромосоме, образуют группы сцепления; обычно они попадают в гамету вместе. Число групп сцепления равно числу пар хромосом, или гаплоидному числу хромосом. Это экспериментально подтверждено у всех хорошо изученных в этом отношении организмов.
Сцепленное наследование. В случае расположения генов в одной хромосоме отмечается их сцепленное наследование – явление совместного наследования генов, локализованных в одной хромосоме. Сцепленное наследование изучал американский генетик Т. Морган на примере наследования признаков у плодовой мушки дрозофилы. (В XX в. дрозофила стала основным объектом генетических исследований.) Были проведены эксперименты по скрещиванию гомозиготных по доминантным аллелям мух (AABB) с серым телом и длинными крыльями и гомозиготных мух по рецессивным аллелям (aabb) с темным телом и редуцированными крыльями.
У гибридов F1 проявились только доминантные признаки – серая окраска тела и длинные крылья (согласно правилу единообразия гибридов первого поколения).
Затем гибриды F1 вновь скрестили с мухами, гомозиготными по рецессивным аллелям (aabb), т. е. провели анализирующее скрещивание.
Полученные результаты отличались от результатов дигибридного скрещивания. Ученые предположили, что пара неаллельных генов, т.е. генов, контролирующих развитие признаков окраски тела и длины крыльев, локализованы в одной хромосоме. В этом случае у дигетерозиготы возможно образование двух типов гамет (AB и ab) и появление двух типов потомков: с серым телом, длинными крыльями (AaBb) и темным телом с редуцированными крыльями (aabb). Причем таких гибридов должно быть поровну (по 50%), а формула расщепления – 1:1. Однако такого соотношения в эксперименте Моргана также не наблюдалось. Результаты анализирующего скрещивания были следующими: 41,5% серых длиннокрылых мух; 41,5% темных с редуцированными крыльями; 8,5% темных длиннокрылых; 8,5% серых с редуцированными крыльями.
Преобладание серых длиннокрылых и темных с редуцированными крыльями мух свидетельствовало о том, что гены A и B и a и b сцеплены, но не абсолютно. На это указывало появление мух с перекомбинированными признаками (серое тело, редуцированные крылья и темное тело, длинные крылья). Перекомбинация генов явилась следствием кроссинговера гомологичных хромосом у некоторых гамет (17 %) во время мейоза (см. § 18). Гаметы с перекомбинированными аллелями получили название кроссоверных гамет. Возникновение потомков с перекомбинироваиными признаками в экспериментах Т. Моргана – свидетельство нарушения сцепления генов.
В ходе исследований ученые убедились в том, что сцепление генов не всегда бывает полным. Оно может быть нарушено кроссинговером, приводящим к возникновению качественно новых (рекомбинантных) хромосом, вследствие чего образуются новые типы гамет. В итоге у потомков появляются новые комбинации признаков, отсутствующие у родительских особей.
Результаты экспериментов с мухой дрозофилой позволили сформулировать закон сцепленного наследования (или закон Моргана): гены, локализованные в одной хромосоме, часто наследуются совместно, образуя группу сцепления.
Хромосомная теория наследственности. Создание хромосомной теории наследственности стало одним из крупнейших теоретических обобщений генетики. Заслуга создания этой теории принадлежит Т. Моргану и сотрудникам его лаборатории К. Бриджесу, А. Стертеванту и Г. Мёллеру.
Согласно хромосомной теории наследственности, хромосомы являются носителями генов и представляют собой материальную основу наследственности. Каждая пара аллелей локализована в паре гомологичных хромосом, причем каждая из гомологичных хромосом несет по одному из пары аллелей.
Конкретные формы генов – вплели занимают одно и то же место – локус – в гомологичных хромосомах.
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Число групп сцепления равно гаплоидному числу хромосом.
Нарушение сцепления – следствие кроссинговера в мейозе. Частота кроссинговера (процент кроссинговера) между двумя неаллельными генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними: чем ближе расположены гены в хромосоме, тем теснее сцепление между ними и тем реже они разделяются при кроссинговере, и наоборот.
Закономерности, открытые научной школой Т. Моргана, были позднее подтверждены многочисленными исследованиями, проведенными на разных объектах.
Основные положения хромосомной теории наследственности:
· Гены расположены в хромосомах линейно.
· Каждый ген в хромосоме занимает определенное место (локус).
· Каждая хромосома представляет собой группу сцепления генов. Число групп сцепления у каждого вида организмов равно гаплоидному числу хромосом.
· Между гомологичными хромосомами может происходить обмен аллельными генами – кроссинговер.
· Расстояние между генами в хромосоме прямо пропорционально проценту кроссинговера. Это означает, что кроссинговер чаще может происходить между более удаленными друг от друга генами и реже– между близко расположенными генами.
Генетические карты. Принцип построения генетических карт (картирование) хромосом разработали Т. Морган и его ученики. В основу этого принципа положено представление о линейном расположении генов по всей длине хромосомы. Генетические карты хромосом представляют собой схему взаимного расположения генов, находящихся в одной группе сцепления. Иными словами, на генетических картах указан порядок расположения генов в хромосоме и относительные расстояния их друг от друга
Генетические карты строитс








