Ренин-ангиотензин-альдостерон. Поддержание стабильной концентрации ионов натрия в крови и объём циркулирующей крови регулируется системой ренин-ангиотензин-альдостерон, которая влияет также на реабсорбцию воды. Уменьшение объёма крови, вызванное потерей натрия, стимулирует группу клеток, расположенных в стенках приносящих артериол - юкстагломерулярный аппарат (ЮГА). В него входят специализированные рецепторные и секреторные клетки. Активация ЮГА приводит к высвобождению из его секреторных клеток протеолитического фермента ренина. Ренин высвобождается из клеток также в ответ на снижение артериального давления крови.
Ренин воздействует на ангиотензиноген (белок a2-глобулиновой фракции), и расщепляет его с образованием декапептида ангиотензина I. Затем другой протеолитический фермент отщепляет от ангиотензина I два концевых аминокислотных остатка с образованием ангиотензина II. Этот октапептид является одним из наиболее активных средств, способствующих сужению кровеносных сосудов, в том числе артериол. В результате увеличивается кровяное давление, снижаются как почечный кровоток, так и клубочковая фильтрация.
Кроме того, ангиотензин II стимулирует секрецию клетками коркового слоя надпочечников гормона альдостерона. Альдостерон - гормон прямого действия - оказывает эффект на дистальный извитой каналец нефрона. Этот гормон индуцирует в клетках-мишенях синтез:
а) белков, участвующих в транспорте Na+ через люминальную поверхность клеточной мембраны;
б) Na+,K+-АТФазы, встраивающейся в контрлюминальную мембрану и участвующей в транспорте Na+ из клеток канальцев в кровь;
в) ферментов митохондрий, например, цитратсинтазы;
г) ферментов, участвующих в образовании фосфолипидов мембран, что облегчает транспорт Na+ в клетки канальца.
Таким образом, альдостерон повышает скорость реабсорбции Na+ из почечных канальцев (за ионами Na+ пассивно следуют ионы Cl—) и в конечном счёте осмотическую реабсорбцию воды, стимулирует активный перенос К+ из плазмы крови в мочу.
84. Половые гормоны - механизм действия, биологическая роль, образование, строение,
Женские половые гормоны (эстрогены). К ним относятся эстрон, эстрадиол и эстриол. Это стероидные гормоны, синтезируемые из холестерола главным образом в яичниках. Секреция эстрогенов регулируется фолликулостимулирующим и лютеинизирующим гормонами гипофиза (см. таблицу 2). Ткани-мишени - тело матки, яичники, маточные трубы, грудные железы. Механизм действия - прямой. Основная биологическая роль эстрогенов заключается в обеспечении репродуктивной функции в организме женщины.

29.5.2. Мужские половые гормоны (андрогены). Главными представителями являются андростерон и тестостерон. Предшественником андрогенов является холестерол, синтезируются они главным образом в семенниках. Регуляция биосинтеза андрогенов осуществляется гонадотропными гормонами (ФСГ и ЛГ). Андрогены - гормоны прямого действия, они способствуют синтезу белка во всех тканях, особенно в мышцах. Биологическая роль андрогенов в мужском организме связана с дифференцировкой и функционированием репродуктивной системы. Распад мужских половых гормонов осуществляется в печени, конечными продуктами распада являются 17-кетостероиды.

Нарушения функций эндокринных желез: гипер- и гипопродукция гормонов. Примеры заболеваний, связанных с дисфункцией эндокринных желез.
(Освещено в предыдущих вопросах)
86. Белки плазмы крови — биологическая роль. Гипо- и гиперпротеинемии, диспротеинемия. Альбумин — функции, причины гипоальбуминемии и ее проявления. Возрастные особенности белковогосостава плазмы крови. Иммуноглобулины. Белки острой фазы. Диагностическое значение определения фракций белков плазмы крови.
Плазма крови содержит сложную многокомпонентную (более 100) смесь белков, различающихся по происхождению и функциям. Большинство белков плазмы синтезируется в печени. Иммуноглобулины и ряд других защитных белков иммунокомпетентными клетками.
Содержание общего белка в сыворотке крови здорового человека составляет 65 - 85 г/л (в плазме крови этот показатель на 2 – 4 г/л выше за счёт фибриногена).
30.2.1. Белковые фракции. При помощи высаливания белков плазмы можно выделить альбуминовую и глобулиновую фракции. В норме соотношение этих фракций составляет 1,5 – 2,5. Использование метода электрофореза на бумаге позволяет выявить 5 белковых фракций (в порядке убывания скорости миграции): альбумины, α1-, α2-, β- и γ-глобулины. При использовании более тонких методов фракционирования в каждой фракции, кроме альбуминовой, можно выделить целый ряд белков (содержание и состав белковых фракций сыворотки крови см. рисунок 1).

Рисунок 1. Электрофореграмма белков сыворотки крови и состав белковых фракций.
Альбумины – белки с молекулярной массой около 70000 Да. Благодаря гидрофильности и высокому содержанию в плазме играют важную роль в поддержании коллоидно-осмотического (онкотического) давления крови и регуляции обмена жидкостей между кровью и тканями. Выполняют транспортную функцию: осуществляют перенос свободных жирных кислот, желчных пигментов, стероидных гормонов, ионов Са2+, многих лекарств. Альбумины также служат богатым и быстро реализуемым резервом аминокислот.
α1-Глобулины:
· Кислый α1-гликопротеин (орозомукоид) – содержит до 40% углеводов, изоэлектрическая точка его находится в кислой среде (2,7). Функция этого белка до конца не установлена; известно, что на ранних стадиях воспалительного процесса орозомукоид способствует образованию коллагеновых волокон в очаге воспаления (Я.Мусил, 1985).
· α1-Антитрипсин – ингибитор ряда протеаз (трипсина, химотрипсина, калликреина, плазмина). Врождённое снижение содержания α1-антитрипсина в крови может быть фактором предрасположенности к бронхо-лёгочным заболеваниям, так как эластические волокна лёгочной ткани особенно чувствительны к действию протеолитических ферментов.
· Ретинолсвязывающий белок осуществляет транспорт жирорастворимого витамина А.
· Тироксинсвязывающий белок – связывает и транспортирует иодсодержащие гормоны щитовидной железы.
· Транскортин – связывает и транспортирует глюкокортикоидные го рмоны (кортизол, кортикостерон).
α2-Глобулины:
· Гаптоглобины (25% α2-глобулинов) – образуют стабильный комплекс с гемоглобином, появляющимся в плазме в результате внутрисосудистого гемолиза эритроцитов. Комплексы гаптоглобин-гемоглобин поглощаются клетками РЭС, где гем и белковые цепи подвергаются распаду, а железо повторно используется для синтеза гемоглобина. Тем самым предотвращается потеря железа организмом и повреждение почек гемоглобином.
· Церулоплазмин – белок, содержащий ионы меди (одна молекула церулоплазмина содержит 6-8 ионов Cu2+), которые придают ему голубую окраску. Является транспортной формой ионов меди в организме. Обладает оксидазной активностью: окисляет Fe2+ в Fe3+, что обеспечивает связывание железа трансферрином. Способен окислять ароматическиеамины, участвует в обмене адреналина, норадреналина, серотонина.
β-Глобулины:
· Трансферрин – главный белок β-глобулиновой фракции, участвует в связывании и транспорте трёхвалентного железа в различные ткани, особенно в кроветворные. Трансферрин регулирует содержание Fe3+ в крови, предотвращает избыточное накопление и потерю с мочой.
· Гемопексин – связывает гем и предотвращает его потерю почками. Комплекс гем-гемопексин улавливается из крови печенью.
· С-реактивный белок (С-РБ) – белок, способный преципитировать (в присутствии Са2+) С-полисахарид клеточной стенки пневмококка. Биологическая роль его определяется способностью активировать фагоцитоз и ингибировать процесс агрегации тромбоцитов. У здоровых людей концентрация С-РБ в плазме ничтожно мала и стандартными методами не определяется. При остром воспалительном процессе она увеличивается более чем в 20 раз, в этом случае С-РБ обнаруживается в крови. Исследование С-РБ имеет преимущество перед другими маркерами воспалительного процесса: определением СОЭ и подсчётом числа лейкоцитов. Данный показатель более чувствителен, его увеличение происходит раньше и после выздоровления быстрее возвращается к норме.
γ-Глобулины:
· Иммуноглобулины (IgA, IgG, IgM, IgD, IgE) представляют собой антитела, вырабатываемые организмом в ответ на введение чужеродных веществ с антигенной активностью. Подробнее об этих белках см. 1.2.5.
30.2.2. Количественные и качественные изменения белкового состава плазмы крови. При различных патологических состояниях белковый состав плазмы крови может изменяться. Основными видами изменений являются:
· Гиперпротеинемия - увеличение содержания общего белка плазмы. Причины: потеря большого количества воды (рвота, диарея, обширные ожоги), инфекционные заболевания (за счёт увеличения количества γ-глобулинов).
· Гипопротеинемия - уменьшение содержания общего белка в плазме. Наблюдается при заболеваниях печени (вследствие нарушения синтеза белков), при заболеваниях почек (вследствие потери белков с мочой), при голодании (вследствие недостатка аминокислот для синтеза белков).
· Диспротеинемия - изменение процентного соотношения белковых фракций при нормальном содержании общего белка в плазме крови, например, снижение содержания альбуминов и увеличение содержания одной или нескольких глобулиновых фракций при различных воспалительных заболеваниях.
· Парапротеинемия - появление в плазме крови патологических иммуноглобулинов - парапротеинов, отличающихся от нормальных белков по физико-химическим свойствам и биологической активности. К таким белкам относятся, например, криоглобулины, образующие друг с другом преципитаты при температуре ниже 37° С. Парапротеины обнаруживаются в крови при макроглобулинемии Вальденстрема, при миеломной болезни (в последнем случае они могут преодолевать почечный барьер и обнаруживаться в моче как белки Бенс-Джонса). Парапротеинемия, как правило, сопровождается гиперпротеинемией.
елки острой фазы воспаления. Это белки, содержание которых увеличивается в плазме крови при остром воспалительном процессе. К ним относятся, например, следующие белки:
1. гаптоглобин;
2. церулоплазмин;
3. С-реактивный белок;
4. α1-антитрипсин;
5. фибриноген (компонент свёртывающей системы крови; см. 30.7.2).
Скорость синтеза этих белков увеличивается прежде всего за счёт снижения образования альбуминов, трансферрина и альбуминов (небольшая фракция белков плазмы, обладающая наибольшей подвижностью при диск-электрофорезе, и которой соответствует полоса на электрофореграмме перед альбуминами), концентрация которых при остром воспалении снижается.
Биологическая роль белков острой фазы: а) все эти белки являются ингибиторами ферментов, освобождаемых при разрушении клеток, и предупреждают вторичное повреждение тканей; б) эти белки обладают иммунодепрессорным действием (В.Л.Доценко, 1985).
30.2.5. Защитные белки плазмы крови. К белкам, выполняющим защитную функцию, относятся иммуноглобулины и интерфероны.
Иммуноглобулины (антитела) - группа белков, вырабатываемых в ответ на попадание в организм чужеродных структур (антигенов). Они синтезируются в лимфоузлах и селезёнке лимфоцитами В. Выделяют 5 классов иммуноглобулинов - IgA, IgG, IgM, IgD, IgE.

Рисунок 3. Схема строения иммуноглобулинов (серым цветом показана вариабельная область, не закрашена - константная область).
Молекулы иммуноглобулинов имеют единый план строения. Структурную единицу иммуноглобулина (мономер) образуют четыре полипептидные цепи, соединённые между собой дисульфидными связями: две тяжёлые (цепи Н) и две лёгкие (цепи L) (см. рисунок 3). IgG, IgD и IgЕ по своей структуре, как правило, являются мономерами, молекулы IgM построены из пяти мономеров, IgA состоят из двух и более структурных единиц, или являются мономерами.
Белковые цепи, входящие в состав иммуноглобулинов, можно условно разделить на специфические домены, или области, имеющие определённые структурные и функциональные особенности.
N-концевые участки как L-, так и Н-цепей называются вариабельной областью (V), так как их структура характеризуется существенными различиями у разных классов антител. Внутри вариабельного домена имеются 3 гипервариабельных участка, отличающихся наибольшим разнообразием аминокислотной последовательности. Именно вариабельная область антител ответственна за связывание антигенов по принципу комплементарности; первичная структура белковых цепей в этой области определяет специфичность антител.
С-концевые домены Н- и L-цепей обладают относительно постоянной первичной структурой в пределах каждого класса антител и называются константной областью (С). Константная область определяет свойства различных классов иммуноглобулинов, их распределение в организме, может принимать участие в запуске механизмов, вызывающих уничтожение антигенов.
Интерфероны - семейство белков, синтезируемых клетками организма в ответ на вирусную инфекцию и обладающих противовирусным эффектом. Различают несколько типов интерферонов, обладающих специфическим спектром действия: лейкоцитарный (α-интерферон), фибробластный (β-интерферон) и& иммунный (γ-интерферон). Интерфероны синтезируются и секретируются одними клетками и проявляют свой эффект, воздействуя на другие клетки, в этом отношении они подобны гормонам. Механизм действия интерферонов показан на рисунке 4.

Рисунок 4. Механизм действия интерферонов (Ю.А.Овчинников, 1987).
Связываясь с клеточными рецепторами, интерфероны индуцируют синтез двух ферментов — 2',5'-олигоаденилатсинтетазы и протеинкиназы, вероятно, за счет инициации транскрипции соответствующих генов. Оба образующихся фермента проявляют свою активность в присутствии двухцепочечных РНК, а именно такие РНК являются продуктами репликации многих вирусов или содержатся в их вирионах. Первый фермент синтезирует 2',5'-олигоаденилаты (из АТФ), которые активируют клеточную рибонуклеазу I; второй фермент фосфорилирует фактор инициации трансляции IF2. Конечным результатом этих процессов является ингибирование биосинтеза белка и размножения вируса в инфицированной клетке (Ю.А.Овчинников, 1987).
87. Низкомолекулярные азотсодержащие вещества крови ("остаточный азот'') и диагностическое значение их определения. Гиперазотемия (ретенционная и продукционная).
К этой группе веществ относятся: мочевина, мочевая кислота, аминокислоты, креатин, креатинин, аммиак, индикан, билирубин и другие соединения (см. рисунок 5). Содержание остаточного азота в плазме крови здоровых людей - 15-25 ммоль/л. Повышение содержания остаточного азота в крови называется азотемией. В зависимости от причины, азотемия подразделяется на ретенционную и продукционную.
Ретенционная азотемия возникает при нарушении выведения продуктов азотистого обмена (в первую очередь мочевины) с мочой и характерна для недостаточности функции почек. В этом случае до 90% небелкового азота крови приходится на азот мочевины вместо 50% в норме.
Продукционная азотемия развивается при избыточном поступлении азотистых веществ в кровь вследствие усиленного распада тканевых белков (длительное голодание, сахарный диабет, тяжёлые ранения и ожоги, инфекционные заболевания).
Определение остаточного азота проводят в в безбелковом фильтрате сыворотки крови. В результате минерализации безбелкового фильтрата при нагревании с концентрированной Н2SO4 азот всех небелковых соединений переходит в форму (NH4)2SO4. Ионы NH4+ определяют с помощью реактива Несслера.
· Мочевина - главный конечный продукт обмена белков в организме человека. Образуется в результате обезвреживания аммиака в печени, выводится из организма почками. Поэтому содержание мочевины в крови снижается при заболеваниях печени и возрастает при почечной недостаточности.
· Аминокислоты - поступают в кровь при всасывании из желудочно-кишечного тракта или являются продуктами распада тканевых белков. В крови здоровых людей среди аминокислот преобладают аланин и глутамин, которые наряду с участием в биосинтезе белков являются транспортными формами аммиака.
· Мочевая кислота - конечный продукт катаболизма пуриновых нуклеотидов. Содержание её в крови возрастает при подагре (в результате усиленного образования) и при нарушениях функции почек (из-за недостаточного выведения).
· Креатин - синтезируется в почках и печени, в мышцах превращается в креатинфосфат - источник энергии для процессов мышечного сокращения. При заболеваниях мышечной системы содержание креатина в крови значительно возрастает.
· Креатинин - конечный продукт азотистого обмена, образуется в результате дефосфорилирования креатинфосфата в мышцах, выводится из организма почками. Содержание креатинина в крови снижается при заболеваниях мышечной системы, повышается при почечной недостаточности.
· Индикан - продукт обезвреживания индола, образуется в печени, выводится почками. Содержание его в крови снижается при заболеваниях печени, повышается - при усилении процессов гниения белков в кишечнике, при заболеваниях почек.
· Билирубин (прямой и непрямой) - продукты катаболизма гемоглобина. Содержание билирубина в крови увеличивается при желтухах: гемолитической (за счёт непрямого билирубина), обтурационной (за счёт прямого билирубина), паренхиматозной (за счёт обеих фракций).
88. Буферные системы крови и кислотно-основное состояние (КОС). Роль дыхательной и выделительной систем в поддержании КОС. Нарушения кислотно-основного баланса. Особенности регуляции КОС у детей .
Буферные системы крови. Буферные системы организма состоят из слабых кислот и их солей с сильными основаниями. Каждая буферная система характеризуется двумя показателями:
· рН буфера (зависит от соотношения компонентов буфера);
· буферная ёмкость, то есть количество сильного основания или кислоты, которое нужно прибавить к буферному раствору для изменения рН на единицу (зависит от абсолютных концентраций компонентов буфера).
Различают следующие буферные системы крови:
· бикарбонатная (H2CO3/NaHCO3);
· фосфатная (NaH2PO4/Na2HPO4);
· гемоглобиновая (дезоксигемоглобин в качестве слабой кислоты/ калиевая соль оксигемоглобина);
· белковая (действие её обусловлено амфотерностью белков). Бикарбонатная и тесно связанная с ней гемоглобиновая буферные системы составляют в совокупности более 80% буферной ёмкости крови.
30.6.2. Дыхательная регуляция КОС осуществляется путём изменения интенсивности внешнего дыхания. При накоплении в крови СО2 и Н+ усиливается лёгочная вентиляция, что приводит к нормализации газового состава крови. Снижение концентрации углекислоты и Н+ вызывает уменьшение лёгочной вентиляции и нормализацию данных показателей.
30.6.3. Почечная регуляция КОС осуществляется главным образом за счёт трёх механизмов:
· реабсорбции бикарбонатов (в клетках почечных канальцев из Н2О и СО2 образуется угольная кислота Н2СО3; она диссоциирует, Н+ выделяется в мочу, НСО3— реабсорбируетоя в кровь);
· реабсорбции Na+ из клубочкового фильтрата в обмен на Н+ (при этом Na2HPO4 в фильтрате переходит в NaH2PO4 и увеличивается кислотность мочи);
· секреции NH4+ (при гидролизе глутамина в клетках канальцев образуется NH3; он взаимодействует с H+, образуются ионы NH4+, которые выводятся с мочой.
30.6.4. Лабораторные показатели КОС крови. Для характеристики КОС используют следующие показатели:
· рН крови;
· парциальное давление СО2 (рСО2) крови;
· парциальное давление О2 (рО2) крови;
· содержание бикарбонатов в крови при данных значениях рН и рСО2 (актуальный или истинный бикарбонат, АВ);
· содержание бикарбонатов в крови пациента в стандартных условиях, т.е. при рСО2=40 мм рт.ст. (стандартный бикарбонат, SB);
· сумма оснований всех буферных систем крови (ВВ);
· избыток или дефицит оснований крови по сравнению с нормальным для данного пациента показателем (BE, от англ. base excess).
Первые три показателя определяются непосредственно в крови с помощью специальных электродов, на основании полученных данных рассчитываются остальные показатели с помощью номограмм или формул.
30.6.5. Нарушения КОС крови. Известны четыре главные формы нарушений кислотно-основного состояния:
· метаболический ацидоз - возникает при сахарном диабете и голодании (за счёт накопления кетоновых тел в крови), при гипоксии (за счёт накопления лактата). При этом нарушении снижается рСО2 и [НСО3-] крови, увеличивается экскреция NH4+ с мочой;
· дыхательный ацидоз - возникает при бронхите, пневмонии, бронхиальной астме (в результате задержки углекислоты в крови). При этом нарушении повышается рСО2 и [HCO3-] крови, увеличивается экскреция NH4+ с мочой;
· метаболический алкалоз - развивается при потере кислот, например, при неукротимой рвоте. При этом нарушении повышается рСО2 и [HCO3-] крови, увеличивается экскреция НСО3- с мочой, снижается кислотность мочи.
· дыхательный алкалоз - наблюдается при усиленной вентиляции лёгких, например, у альпинистов на большой высоте. При этом нарушении снижается рСО2 и [НСО3-] крови, уменьшается кислотность мочи.
Для лечения метаболического ацидоза используют введение раствора бикарбоната натрия; для лечения метаболического алкалоза - введение раствора глутаминовой кислоты.
89. Метаболизм эритроцита: роль гликолиза и пентозофосфатного пути. Метгемоглобинемия. Ферментативная антиоксидантная система клетки. Причины и последствия дефицита глюкозо-6-фосфатдегидрогеназы эритроцитов.
Эритроциты - высокоспециализированные клетки, основной функцией которых является транспорт кислорода из лёгких в ткани. Продолжительность жизни эритроцитов составляет в среднем 120 суток; разрушение их происходит в клетках ретикуло-эндотелиальной системы. В отличие от большинства клеток организма, у эритроцита отсутствуют клеточное ядро, рибосомы и митохондрии.
30.8.2. Энергетический обмен. Основным энергетическим субстратом эритроцита является глюкоза, которая поступает из плазмы крови путём облегчённой диффузии. Около 90% используемой эритроцитом глюкозы подвергается гликолизу (анаэробному окислению) с образованием конечного продукта - молочной кислоты (лактата). Запомните функции, которые выполняет гликолиз в зрелых эритроцитах:
1) в реакциях гликолиза образуется АТФ путём субстратного фосфорилирования. Основное направление использования АТФ в эритроцитах - обеспечение работы Na+,K+-АТФазы. Этот фермент осуществляет транспорт ионов Nа+ из эритроцитов в плазму крови, препятствует накоплению Na+ в эритроцитах и способствует сохранению геометрической формы этих клеток крови (двояковогнутый диск).
2) в реакции дегидрирования глицеральдегид-3-фосфата в гликолизе образуется НАДН. Этот кофермент является кофактором фермента метгемоглобинредуктазы, участвующей в восстановлении метгемоглобина в гемоглобин по следующей схеме:

Эта реакция препятствует накоплению метгемоглобина в эритроцитах.
3) метаболит гликолиза 1,3-дифосфоглицерат способен при участии фермента дифосфоглицератмутазы в присутствии 3-фосфоглицерата превращаться в 2,3-дифосфоглицерат:
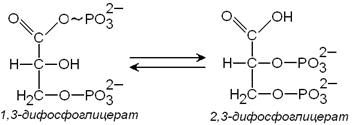
2,3-Дифосфоглицерат принимает участие в регуляции сродства гемоглобина к кислороду. Его содержание в эритроцитах повышается при гипоксии. Гидролиз 2,3-дифосфоглицерата катализирует фермент дифосфоглицератфосфатаза.
Приблизительно 10% глюкозы, потребляемой эритроцитом, используется в пентозофосфатном пути окисления. Реакции этого пути служат основным источником НАДФН для эритроцита. Данный кофермент необходим для перевода окисленного глутатиона (см. 30.8.3) в восстановленную форму. Дефицит ключевого фермента пентозофосфатного пути - глюкозо-6-фосфатдегидрогеназы - сопровождается уменьшением в эритроцитах отношения НАДФН/НАДФ+, увеличением содержания окисленной формы глутатиона и снижением резиcтентности клеток (гемолитическая анемия).
30.8.3. Механизмы обезвреживания активных форм кислорода в эритроцитах. Молекулярный кислород в определённых условиях может превращаться в активные формы, к которым относятся супероксидный анион О2-, пероксид водорода Н2О2, гидроксильный радикал ОН. и синглетный кислород 1О2. Эти формы кислорода обладают высокой реакционной способностью, могут оказывать повреждающее действие на белки и липиды биологических мембран, вызывать разрушение клеток. Чем выше содержание О2, тем больше образуется его активных форм. Поэтому эритроциты, постоянно взаимодействующие с кислородом, содержат эффективные антиоксидантные системы, способные обезвреживать активные метаболиты кислорода.
Важным компонентом антиоксидантных систем является трипептид глутатион, образующийся в эритроцитах в результате взаимодействия γ-глутамилцистеина и глицина:

Восстановленная форма глутатиона (сокращённое обозначение Г-SH) участвует в реакциях обезвреживания пероксида водорода и органических пероксидов (R-O-OH). При этом образуются вода и окисленный глутатион (сокращённое обозначение Г-S-S-Г).

Превращение окисленного глутатиона в восстановленный катализирует фермент глутатионредуктаза. Источник водорода - НАДФН (из пентозофосфатного пути, см. 30.8.2):

В эритроцитах имеются также ферменты супероксиддисмутаза и каталаза, осуществляющие следующие превращения:

Антиоксидантные системы имеют для эритроцитов особое значение, так как в эритроцитах не происходит обновления белков путём синтеза.








