Понятие тотипотентности.
Тотипотентность – способность образовывать из клетки организм. Понятие тотипотентности, мультипотентности и плюрипотентности тесно связано с понятиями «потенция к развитию» и «детерминация».
Cтволовые клетки классифицированы в соответствии со своей возможностью к дифференцировке как тотипотентные, плюрипотентные и мультипотентные.
Тотипотентные - клетки, способные дифференцироваться в любые клетки организма. Как из одной оплодотворенной клетки вырастает целый организм.
Плюрипотентные - клетки, способные образовывать множество различных клеток, но не целый организм.
Мультипотентные - клетки, способные образовывать клетки тканей, из которых они были взяты.
Унипотентные - клетки, дающие начало только одному типу клеток.
Клетки развивающегося эмбриона изначально тотипотентны, но теряют это свойство после нескольких клеточных делений, т.е. они дифференцируются. Некоторые из клеток организма не дифференцируются окончательно, а становятся плюрипотентными, т.е. способны давать лишь некоторые типы клеток целого организма. Тотипотентные клетки эмбриона называют также эмбриональными стволовыми клетками (ESC - Embryonic Stem Cells), а плюри- и мультипотентные клетки организма называют взрослыми стволовыми клетками (ASC - Adult Stem Cells). Функция ESC в организме очевидна: из одной клетки должен развиться целый организм с огромным числом клеточных типов (~200 у человека), каждый из которых выполняет свою функцию. ASC необходимы организму для восполнения погибших клеток в процессе жизни. ASC способны заменять практически все ткани в организме: мозг, костный мозг, кровь, почку, эпителий пищеварительной системы, кожу, сетчатку, мышцы, поджелудочную железу и печень.
ASC способны к самоподдержанию и производству клеток-предшественников, которые затем дифференцируются.
Оплодотворённые яйцеклетки животных являются тотипотентными. В течение развития потомки зиготы утрачивают тотипотентность. У большинства животных бластомеры сохраняют тотипотентность в течение определённого количества делений дробления. У животных с детерминированным (т.е. предопределённым) развитием (например, у круглых червей) первые потомки зиготы уже утрачивают тотипотентность: рост организма происходит не за счёт увеличения числа клеток, а за счёт их увеличения в размере.
У некоторых организмов клетки могут дедифференцироваться и обретать тотипотентность. Срезанные части растений и каллус (недифференцированные клетки, являющиеся тотипотентными, способными дать начало целому растению) могут быть использованы для выращивания целого растения.
Отличия прокариот от эукариот:
1. Отсутствие оформленного ядра (ядерной оболочки). Наследственный материал в виде нуклеоида.
2. Кольцевая хромосома.
3. Хромосома одна или же несколько одинаковых.
4. Сопряжение транскрипции и трансляции во времени и пространстве.
5. Нет интронов и, следственно, нет процессинга мРНК.
Интроны - транскрибируемый, но не кодирующий белок участок гена, расположенный между экзонами, который удаляется из состава первичного транскрипта (преРНК) при сплайсинге.
Экзоны - кодирующие участки ДНК (копии которых составляют зрелую РНК).
Сплайсинг - процесс вырезания определённых нуклеотидных последовательностей из молекул РНК и соединения последовательностей, сохраняющихся в "зрелой" молекуле.
Процессинг (посттранскрипционные модификации РНК) - совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК.
6. Рибосомы 70S=50S+30S. А у эукариотов 80S=60S+40S. S - константа седиментации (осаждения в центрифуге).
7. Нет двумембранных органоидов.
8. Нет цитоскелета.
9. Клеточная стенка из муреина (пептидогликана).
Прокариоты делятся на грамположительные (Гр+, краска застревает) и грамотрицательные (Гр-, краска вымывается) по физическим и химическим свойствам клеточной стенки (окрашивание по Грану).
10. Особое строение жгутика.
11. мРНК полицистронная, т.е. кодирующая последовательности более чем одного белка. У эукариот - моноцистронная, т.е. кодирующая одну полипептидную цепь.
12. Одна точка начала репликации - origin => монорепликонная хромосома.
СИМПЛАСТ
Симпласты (от греч. syn - вместе, plastos - вылепленный) – крупные образования, состоящие из цитоплазмы со множеством ядер. Примеры: мышечные волокна, наружный слой трофобласта плаценты (трофобласт обеспечивает контакт зародыша с материнским организмом). Они возникают вторично за счет слияния отдельных клеток или же в результате деления одних ядер без разделения цитоплазмы, без цитотомии. У растений симпласт - это группа клеток, которые связаны между собой посредством плазмодесм, или же слившиеся клетки.
Одно время возражения учёных касались того, что часто у животных кроме отдельных клеток встречаются так называемые симпласты и синцитии (соклетия), а у растительных - плазмодии. По морфологическому описанию - это крупные цитоплазматические образования со множеством ядер, не разделённые на отдельные клеточные территории. Примерами таких симпластов могут быть мышечные волокна позвоночных или эпидермис у ленточных червей, а также плазмодии у низших грибов миксомицетов. Однако если проследить за развитием таких "неклеточных" форм, то легко убедиться в том, что они возникают вторично за счёт слияния отдельных клеток или же в результате деления одних ядер без цитотомии (разделения цитоплазмы).
РЕАКЦИЯ ФЁЛЬГЕНА
Фёльгена реакция, способ гистохимического выявления дезоксирибонуклеиновой кислоты (ДНК) в клетках животных и растительных организмов, а также у бактерий. Ф. р. предложена в 1924 нем. химиком Р. Фёльгеном (R. Feulgen). Состоит из двух этапов. Сначала проводят кислотный гидролиз (обычно 8–12 мин),в результате которого отщепляются азотистые основания и образуются альдегидные группы. Затем препараты помещают в светло-желтый реактив Шиффа (фуксинсернистая кислота), связывающий эти группы. По образующемуся при таком взаимодействии красно-фиолетовому продукту реакции судят о наличии ДНК. Ф. р. возможна после применения любого фиксатора (кроме жидкости Буэна, содержащей пикриновую кислоту). Ф. р. используют для количественного определения ДНК (см.Цитофотометрия). Имеются разные модификации Ф. р. для выявления локализации и структуры ДНК на электронно-микроскопическом уровне.
ПЕРОКСИСОМЫ (МИКРОТЕЛЬЦА)
Пероксисомы - небольшие (0,3-1,5 мкм) одномембранные вакуоли с нуклеоидом в центре (нуклеоид не такой, как в прокариотическом ядре, да и вообще ничего общего с ядерными структурами не имеет). Изолированные сердцевины пероксисом содержат фермент уратоксидазу. Пероксисомы обнаружены у простейших (амёбы), низших грибов (дрожжи), в нек. эмбриональных тканях высших растений (эндосперм), а также в их зелёных частях, способных к фотореспирации (дыханию, индуцируемому светом), у позвоночных (главным образом в печени и почках). Пероксисомы тесно связаны с ЭПР (вероятно, они образовались в результате заполнение плотным материалом расширенных концов цистерн ЭПР), а зелёных растений - с митохондриями и пластидами. Ферменты пероксисом: оксидазы, уратоксидаза, оксидаза d-аминокислот - при их работе образуется H2O2 и каталаза, разрушающая её (т.е. каталаза выполняет защитную функцию, т.к. перекись водорода токсична). В пероксисомах цыплят и лягушек кроме уратоксидазы содержатся фермента катаболизма пуринов. У животных и нек. растений пероксисомы участвуют в превращении жиров в углеводы.
СФЕРОСОМЫ
Сферосомы - мембранные пузырьки (d 100-150 нм) в клетках растений, окрашивающиеся липофильными красителями и имеющие высокий коэффициент преломления (поэтому хорошо видны в световой микроскоп). Сферосомы образуются в рез-те накопления на конце цистерны ЭПР осмиофильного материала и отшнуровывания этого участка.
ВАКУОЛИ РАСТИТЕЛЬНЫХ КЛЕТОК
У молодых клеток может быть несколько маленьких вакуолей, которые с возрастом сливаются, образуя одну или несколько крупных вакуолей, занимающих до 80% объёма всей клетки. Тонопласт - мембрана, отграничивающая центральную вакуоль от цитоплазмы. Центральная вакуоль образуется из отщепившихся от ЭПР мелких пузырьков, которые увеличиваются в объёме и сливаются друг с другом. Полость вакуоли заполнена клеточным соком - водным раствором с растворёнными молекулами.
Функции центральной вакуоли:
1. Поддержание тургорного давления. Тургорное давление - внутреннее давление, которое развивается в растительной клетки в рез-те поступления воды при осмосе. Цитоплазма прижимается к клеточной стенке, что мешает проникновению в клетку воды. Растворённые в клеточном соке молекулы определяют осмотическую концентрацию. Вакуоль функционирует в качестве осмометра клетки и определяет необходимую прочность и тургисцентность (напряжённость).
2. Накопление запасных веществ (гликозидов - нерастворимых в воде орг. компонентов, соединённых с молекулами сахаров - к примеру, антоцианы).
3. Выброс метаболитов - экскреция: алкалоиды (никотин, кофеин), полифенолы.
______
Вакуолярные структуры способны переходить друг в друга при перестройке мембран.
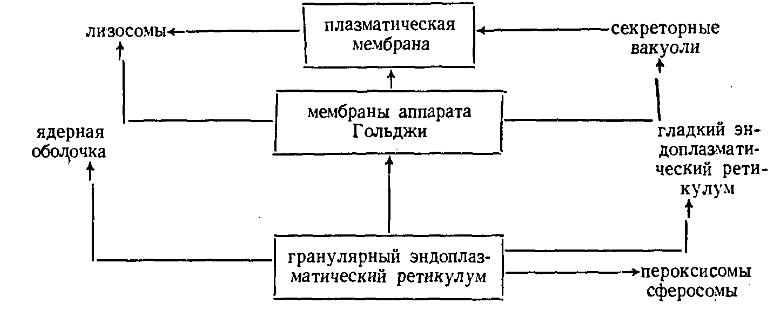
Обратного генезиса мембран ЭПР из мембран АГ нет, т.к. первичная сборка мембранных элементов идёт в грЭПР, а их доп. специализация связана с мембранами АГ.
По оценкам учёных, сегодня на Земле существует более 10 млн., а возможно, даже 100 млн. видов организмов.
Наследственность отличает жизнь от других процессов типа роста кристалла, или горения свечи, или формирования волн на воде, в ходе которых производятся упорядоченные структуры, но единообразного типа связи между особенностями родителей и потомка нет.
Двухцепочные (двунитевые) молекулы ДНК - длинные неразветвлённые спаренные полимерные цепи, всегда образованные из одних и тех же мономеров: A (аденин), T (тимин), C (цитозин), G (гуанин). Мы можем взять часть ДНК из клетки человека и вставить её в бактерию или наоборот - в обоих случаях информация будет успешно читаться, интерпретироваться и копироваться. С помощью химических методов учёные могут прочитать полную последовательность мономеров в любой молекуле ДНК и таким образом расшифровать наследственную информацию, которую содержит любой организм.
КЛЕТОЧНАЯ ТЕОРИЯ
Клеточная теория - это обобщённые представления о строении клеток как единиц живого, об их размножении и роли в формировании многоклеточных организмов.
Появлению и формулированию отдельных положений клеточной теории предшествовал длительный период накопления наблюдений над строением различных одноклеточных и многоклеточных организмов растений и животных. Этот период связан с развитием оптических методов исследования.
1665 - Роберт Гук наблюдал с помощью увеличительных линз подразделение тканей пробки на "ячейки", или "клетки". Его описания послужили толчком для появления систематических исследований анатомии растений (Мальпиги, 1671; Грю, 1671).
1680 - Антони ван Левенгук открыл мир одноклеточных организмов и впервые увидел клетки животных (эритроциты). Позднее клетки животных были описаны Ф. Фонтана.
Прогресс в изучении микроанотомии и клетки связан с развитием микроскопирования в XIX веке. Главным в организации клетки стала считаться не клеточная стенка, а протоплазма.
1833 - Браун открыл ядро.
Все эти многочисленные наблюдения позволили Теодору Шванну в 1838 г. сделать ряд обобщений. Он показал, что клетки животных и растений гомологичны, то есть принципиально сходны между собой.
Гомологичность клеток
Термин "гомологичность" означает сходство по коренным свойствам и отличие по второстепенным. Это обобщение сделано Т. Шванном. Гомологичность строения клеток наблюдается внутри каждого из типов клеток: прокариотическом и эукариотическом. Такое одновременное сходство строения и разнообразие форм определяются тем, что клеточные функции можно грубо подразделить на обязательные и факультативные. Обязательные - направленные на поддержание жизнеспособности самих клеток - осуществляются специальными внутриклеточными структурами (пример: плазмалемма). Одновременно видно разнообразие клеток и внутри одного многоклеточного организма (различия между клетками разных тканей). Другими словами, гомологичность в строении клеток определяется сходством общеклеточных функций, направленных на поддержание жизни самих клеток и на их размножение. Разнообразие же - результат функциональной специализации или следствие эволюционной приспособленности, изменчивости.
Клетка от клетки
Р. Вирхов: "Omnis cellula e cellula".
Шванн подчёркивал одинаковость принципа развития животных и растительных клеток. Это представление базировалось на выводах Шлейдена о том, что клетки могут образовываться из зернистой массы в недрах клеток заново (теория цитобластемы). Вирхов как противник идеи о самозарождении жизни настаивал на "преемственном размножении клеток". Размножение клеток прокариотических и эукариотических происходит только путём деления исходной клетки, которому предшествует воспроизведение её ген. материала (редупликация ДНК). Прокариотические клетки обычно делятся бинарным образом, простой перегородкой, без участия специальных аппаратов деления. У эукариотических клеток - митоз (или мейоз при образовании половых клеток): образуется специальный аппарат клеточного деления - клеточное веретено, с помощью которого хромосомы равномерно распределяются по дочерним клеткам. Другая форма клеточных делений - амитоз, или простое деление - может встречаться в ряде патологических случаев или при делении полиплоидных ядер.
Клетки и организм
Шванн представлял себе многогранную деятельность организма как сумму жизнедеятельности отдельных клеток. Это представление было расширено Вирховым (теория "клеточного государства"). Действительно, какую бы сторону деятельности целого организма мы ни брали, каждая из них осуществляется специализированными клетками. Клетка - единица функционирования в многоклеточном организме. Клетки объединены в функциональные системы: ткани и органы, взаимосвязанные между собой. Вот почему мы говорим об организме как о целом. Специализация частей многоклеточного единого организма, расчленённость его функций дают ему большие возможности приспособления для сохранения вида.
Предположение Вирхова (1858): клетки как единицы развития патологических процессов в организме служат в момент всех заболеваний без исключения мишенью нарушения обменных или регуляторных процессов в конкретных клетках. К примеру, сахарный диабет. Причина его возникновения - недостаточность функционирования лишь одной группы клеток, так называемых B-клеток островков Лангерганса в поджелудочной железе. Эти клетки вырабатывают гормон инсулин, участвующий в регуляции сахарного обмена организма.
МЕТОДЫ ЦИТОЛОГИИ
Цитология возникла как ветвь микроанатомии, и поэтому основной метод, использующийся в цитологии, - метод световой микроскопии. С введением электронной микроскопии в ряде случаев уже трудно провести границу между собственно цитологией и биохимией, они объединяются на уровне макромолекулярного изучения объектов (например, мембран, микротрубочек, микрофиламентов). Всё же главным методическим приёмом в цитологии остаётся визуальное наблюдение объекта - глазом, вооружённым увеличивающими оптическими системами. Кроме того, в цитологии применяются многочисленные приёмы препаративной и аналитической биохимии, методы биофизики.
Световая микроскопия
Микроскопический метод (гр. micros - мельчайший, scopeo - смотрю) позволяет изучать структуру клетки с помощью микроскопов (светового, фазово-контрастного, люминесцентного, ультрафиолетового, электронного). При световой микроскопии объект рассматривается в лучах видимого света. Для этого используются микроскопа типа МБР, МБИ, МБС-1, МИКМЕД-1 и др.
Разрешающая способность микроскопа даёт раздельное изображение двух близких друг к другу линий. Невооружённый человеческий глаз имеет разрешающую способность около 0,1 мм, или 100 мкм. Лучший световой микроскоп примерно в 500 раз улучшает возможность человеческого глаза, т.е. его разрешающая способность составляет около 0,2 мкм, или 200 нм.
Разрешающая способность и увеличение не одно и то же. Различают полезное и бесполезное увеличения. Под полезным понимают такое увеличение наблюдаемого объекта, при котором можно выявить новые детали его строения. Бесполезное увеличение - при котором при увеличении объекта нельзя обнаружить новые детали строения.
Изображения, даваемые объективом, можно увеличить во много раз, применяя сильный окуляр или, например, путём проекции на экран (до 105 раз). Вычислено, что разрешающая способность объектива, т.е. минимальное расстояние между двумя точками, которые видны раздельно, будет равно
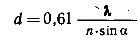
где λ - длина волны света, используемого для освещения объекта
n - коэффициент преломления среды
α - угол между оптической осью объектива и наиболее отклоняющимся лучом, попадающим в объектив
Разрешение микроскопа зависит от длины волны: чем она меньше, тем меньшего размера деталь мы можем увидеть, и от нумерической апертуры объектива ( n * sinα ): чем она выше, тем выше разрешение. Обычно в световых микроскопах используются источники освещения в видимой области спектра (400-700 нм), поэтому максимальное разрешение микроскопа в этом случае может быть не выше 200-350 нм (0,2-0,35 мкм). Если использовать ультрафиолетовый свет (260-280 нм), то можно повысить разрешение до 130-140 нм (0,13-0,14 мкм). Это будет пределом теоретического разрешения светового микроскопа, определяемого волновой природой света. Таким образом, всё, что может дать световой микроскоп как вспомогательный прибор к нашему глазу, - это повысить разрешающую способность его примерно в 1000 раз (невооружённый глаз человека имеет разрешающую способность около 0,1 мм, или 100 мкм). Это и есть "полезное" увеличение микроскопа, выше которого можно увеличивать только контуры изображения, не открывая в нём новых деталей. Следовательно, при использовании видимой области света 0,2-0,3 мкм является конечным пределом разрешения светового микроскопа.
Но всё же в световом микроскопе можно видеть частицы меньшей величины, чем 0,2 мкм. Это метод "тёмного поля", или, как его называли раньше, метод "ультрамикроскопии". Суть его в том, что подобно пылинкам в луче света (эффект Тиндаля) в клетке при боковом освещении светятся мельчайшие частицы (меньше 0,2 мкм), отражённый свет от которых попадает в объектив микроскопа. Этот метод успешно применяется при изучении живых клеток.
Большая часть клеточных компонентов мало отличается по свойствам от среды и друг от друга и поэтому мало заметны и не контрастны. Для их изучения приходится изменять освещённость (теряя при этом в чёткости изображения) или применять особые методы и приборы.
Метод фазово-контрастной микроскопии основан на том, что отдельные участки прозрачной клетки мало, но всё же отличаются друг от друга по плотности и светопреломлению. В фазово-контрастном микроскопе в объектив вмонтирована специальная пластинка, проходя через которую луч света испытывает дополнительный сдвиг фазы колебаний. Создаётся светло-тёмное контрастное изображение объекта.
Сходный приём используется в интерференционном микроскопе. Он устроен так, что пучок параллельных световых лучей от осветителя разделяется на два потока. Один из них проходит через объект и приобретает изменения в фазе колебания, а другой идёт, минуя объект. В призмах объектива оба потока вновь соединяются и интерферируют между собой. В результате интерференции будет строиться изображение, на котором участки клетки, обладающие разной толщиной или разной плотностью, будут отличаться друг от друга по степени контрастности.
С помощью поляризационного микроскопа изучают объекты, обладающие так называемой изотропией, т.е. упорядоченной ориентацией субмикроскопических частиц (например, волокна веретена деления, миофибриллы и др.). У такого микроскопа перед конденсором помещается поляризатор, пропускающий световые волны с определённой плоскостью поляризации. После препарата и объектива помещается анализатор, пропускающий свет с этой же плоскостью поляризации. Поляризатор и анализатор - это призмы. С помощью поляризационного микроскопа можно убедиться, например, в ориентированном расположении мицелл в клеточной стенке растений.
http://snablab.ru/stati/metody-mikroskopirovaniya
Микрохирургия.
С помощью микроманипулятора клетки разрезают, извлекают из них части, вводят вещества (микроинъекция) и т.д. За ходом операции наблюдают через обычный микроскоп, совмещённый с микроманипулятором. Микрохирургические инструменты: стеклянные крючки, иглы, капилляры, которые изготавливаются на "микрокузницах". При микроманипуляции клетки помещают в специальные камеры, в которые вводят инструменты. Так, с помощью микроманипулятора удалось пересадить ядра от одного штамма амёбы другому и доказать, что именно клеточное ядро определяет физиологические особенности клетки в целом. Также удалось инъецировать в клетку амёбы коллоидное золото и исследовать распределение его частиц в цитоплазме и ядре. С помощью микрохирургических инструментов можно поворачивать в клетках митотические веретена, оттаскивать отдельные хромосомы и т.д. Также применяют микропучки у/ф света или лазерные микропучки.
Флуоресцентная микроскопия:
Ряд веществ способен светиться (флуоресцировать, люминесцировать) при поглощении ими световой энергии. Спектр флуоресценции всегда смещён в сторону больших длин волн по отношению к возбуждающему флуоресценцию излучению. Так, например, выделенный хлорофилл при освещении в у/ф лучах светится красным цветом. Этот принцип используется в флуоресцентной микроскопии: рассматривание флуоресцирующих объектов в зоне коротких длин волн. Обычно в таких микроскопах используются фильтры, дающие освещение в сине-фиолетовой области.
Собственной флуоресценцией обладают некоторые пигменты (хлорофиллы, бактериальные пигменты), витамины (А, B2), гормоны. Можно применять метод флуоресцентной микроскопии, добавляя к живым клеткам флуорохромы (флуоресцирующие вещества).
ЭЛЕКТРОННАЯ МИКРОСКОПИЯ
Ультрамикротомия
При прохождении пучка электронов через объект часть электронов поглощается, что приводит к нагреванию объекта и его деформации, поэтому необходимо иметь тонкие объекты (не выше 0,1 мкм). Но даже если мы будем рассматривать неизменяющиеся объекты большой толщины (0,5-1 мкм), на конечном изображении будут наслаиваться проекции структур, располагающихся на разных уровнях по толщине объекта.
Процедура создания ультратонких срезов в электронной микроскопии сходна с той, что используется в световой микроскопии: фиксация клеток и тканей (фиксаторы - буферные растворы глютарового альдегида или четырёхоксиси осмия; или же двойная фиксация: сначала глютаровый альдегид, затем осмий); обезвоживание; пропитывание эпоксидными смолами или другими пластиками; полимеризация таких пластмасс. Для разрезания используют алмазные ножи (режущая поверхность сколов стекла идеально острая и без зазубрин, но стеклянные ножи используют только один раз ввиду их недолговечности). Площадь получаемых ультратонких срезов обычно 0,1-1мм2, поэтому все операции проходят под микроскопическим контролем. Срезы, смонтированные на сетках с подложкой, необходимо дополнительно контрастировать - "окрашивать" при помощи солей тяжёлых металлов: свинца, урана.
Методы приготовления ультратонких срезов без фиксации и заливки клеток в твёрдые пластмассы - методы криоультрамикротомии, т.е. получение срезов с замороженных тканей, моментально охлаждённых до температуры жидкого азота (-196 градусов). При этом происходит практически одномоментное торможение всех метаболических процессов, а вода из жидкой фазы переходит в твёрдую, но не кристаллическую: её молекулярная структура беспорядочна (стекловидное состояние). Такие твёрдые блоки можно резать при температуре жидкого азота на ультратонкие срезы (нож при этом также охлаждён).
Фракцирование клеток
В настоящее время в виде чистых фракций получают почти любые клеточные органоиды и структуры: ядра, ядрышки, хроматин, плазматическую мембрану, вакуоли ЭПР, аппарат Гольджи, пероксисомы, митохондрии и т.д.
Получение клеточных фракций: общее разрушение клетки, гомогенезация (доведение до однородной структуры путём разрушения клеточных границ, главным образом механическими способами: ультразвуковая дезинтеграция, растирание в смеси со стеклянными шариками, пропускание через узкие отверстия, дифференциальное (разделительное) центрифугирование (чем больше, плотнее, тяжелее частица, тем быстрее она осядет на дно пробирки + ускорения центрифуги. При повторном центрифугировании этих смешанных подфракций можно получить чистые фракции); при этом не должны разрушаться внутриклеточные компоненты).
ДНК Хроматина
Примерно 30-40%. Двухцепочная спиральная молекула. Гиперхромный эффект: повышение оптической плотности раствора при нагревании растворов хроматина; связан с разрывом межнуклеотидных водородных связей между цепями ДНК. Длина молекул ДНК не изменятся после обработки препаратов протеолитическими ферментами. У микроорганизмов кол-во ДНК на клетку значительно ниже, чем у высших растений и животных (так, у мыши на ядро приходится почти в 600 раз больше ДНК, чем у кишечной палочки). Сравнивая кол-во ДНК на клетку у эукариотов, трудно уловить какие-либо корреляции между степенью сложности организма и количеством ДНК на ядро (одно и то же кол-во ДНК на клетку может быть у совершенно разных организмов; у более просто устроенного организма это кол-во может быть больше, чем у сложно устроенного). Значительны колебания кол-ва ДНК в больших таксономических группах.
Как решить этот вопрос? Если фрагментированные молекулы ДНК в растворах подвергнуть тепловой денатурации, а затем инкубировать их при температуре ниже той, при которой происходила денатурация, идёт восстановление исходной двуспиральной структуры фрагментов ДНК за счёт воссоединения комплементарных цепей - ренатурация. обратимая и необратимая денатурация. Для ДНК вирусов и прокариотов было показано, что скорость ренатурации прямо зависит от величины генома: чем больше геном, чем больше кол-во ДНК на частицу или клетку, тем больше нужно времени для случайного сближения комплементарных цепей. Это указывает на отсутствие повторяющихся последовательностей оснований в геноме прокариотов; все участки их ДНК несут уникальные последовательности.
В состав же эукариотических ДНК входят фракции, ренатурирующие с гораздо большей скоростью, чем можно было бы предполагать на основании размера их генома, а также фракция ДНК, ренатурирующая медленно, подобно уникальным последовательностям ДНК прокариотов. Однако для эукариотов требуется значительно большее время для ренатурации этой фракции, что связано с общим большим размером их генома и с большим числом уникальных генов.
!!!!!!!!!!В части ДНК эукариотов с высокой скоростью ренатурации различают две подфракции: 1. с высоко или часто повторяющимися последовательностями (участки ДНК могут быть повторены 106 раз; 2. умеренно повторяющихся последовательностей (102-103 раз). Фракции с часто повторяющимися последовательностями могут обладать иной плавучей плотностью, чем основная масса ДНК, и поэтому могут быть выделены в чистом виде, как так называемая фракция сателлитной ДНК. Сателлитная ДНК не участвует в синтезе основных типов РНК, не связана с процессом синтеза белка, т.к. ни один из типов РНК не гибридизируется с сателлитными ДНК, т.е. сателлитные ДНК не являются матрицами для синтеза РНК, не участвуют в транскрипции. Итак, ДНК эукариотических клеток гетерогенна по составу, содержит несколько классов последовательностей нуклеотидов: часто повторяющиеся последовательности (≥106 раз), входящие во фракцию сателлитной ДНК и не транскрибирующиеся; фракции умеренно повторяющихся последовательностей (102-105), представляющих блоки истинных генов, а также короткие последовательности, разбросанные по всему геному; фракция уникальных последовательностей, несущая информацию для большинства белков клетки.
При использовании авторадиографического метода исследования молекул ДНК было найдено, что ДНК эукариотических хромосом представляет собой линейные молекулы, состоящие из тандемно (друг за другом) расположенных репликонов (единиц репликации - участков ДНК, синтезирующихся как независимые единицы) разного размера. Средний размер репликона около 30 мкм. Как и в случае прокариотических клеток, эти репликоны имеют начальную и терминальную точки синтеза ДНК. Благодаря полирепликонному строению молекул ДНК весь процесс репликации занимает 7-12 ч. В этот период происходит синтез всей ядерной ДНК (синтетический, или S-период). Однако синтез ДНК среди хромосом происходит асинхронно: начинается и кончается не во всех хромосомах одновременно. Длительность процесса репликации ДНК в отдельных хромосомах не зависит от их размера. Это явление отображает межхромосомную асинхронность синтеза ДНК.
Внутрихромосомная асинхронность: различные участки одной хромосомы редуплицируются не одновременно. Наиболее поздно репликация заканчивается в хромосомах или их участках, находящихся в конденсированном состоянии.
Тейлор: редупликация хромосом аналогична полуконсервативной редупликации ДНК, т.е. хромосомная ДНК воспроизводится так, как если бы в составе хромосом была одна молекула ДНК.
Белки хроматина
Фракция белков хроматина - 60-70% сухой массы. Белки хроматина: гистоны и негистоновые белки. В валовом отношении негистоновые белки составляют 20% от кол-ва гистонов.
Гистоны - сильноосновные белки, экстрагируемые (выделяемые) из очищенного хроматина с помощью кислых растворов или крепких растворов солей. Щелочность гистонов связана с их обогащённостью основными аминокислотами (главным образом лизином и аргинином). Гистоны не содержат триптофана.
Препарат суммарных гистонов можно разделить на 5 фракций:
Н1 (от англ. histone) - богатый лизином гистон
Н2б - умеренно богатый лизином гистон
Н2а - умеренно богатый лизином гистон
Н4 - богатый аргинином гистон
Н3 - богатый аргинином гистон
Эти пять фракций гистонов присутствуют практически во всех клетках растений, рыб, рептилий, птиц, млекопитающих. В препаратах хроматина эти фракции гистонов обнаруживаются в приблизительно равных кол-вах, кроме Н1, которого примерно в 2 раза меньше любой из других фракций. Гистоны, кроме Н1, сходны по своей первичной структуре (по последовательности а/к в полипептидной цепи). Гистон Н1 наиболее вариабелен по сравнению с другими гистонами, может включать в себя 3-6 субфракций.
Для молекул гистонов характерно неравномерное распределение основных а/к вдоль цепи: обогащение положительно заряженными аминогруппами наблюдается на концах белковых цепей. Видимо, эти участки гистонов связываются с фосфатными группировками на ДНК. Таким образом, взаимодействие между гистонами и ДНК, приводящее к образованию дезоксирибонуклеопротеидного комплекса, носит ионный характер.
Протамины - богатые аргинином белки.
Прокариотические клетки не содержат связанных с ДНК сильноосновных белков, аналогичных гистонам.
Все пять гистонов в ядрах подвергаются посттрансляционным изменениям а/к. Эти модификации могут сказываться на структуре хроматина, приводить к изменению его синтетической активности.
Гистоны синтезируются на полисомах в цитоплазме. Этот синтез начинается несколько раньше редупликации ДНК.
Функциональная роль гистонов не вполне ясна. Одно время считалось, что гистоны являются специфическими регуляторами активности ДНК хроматина, но одинаковость строения гистонов не только в разных клетках одного организма, но и в клетках разных организмов, говорит о малой вероятности этого. Более очевидна структурная роль гистонов: обеспечение специфической укладки хромосомной ДНК, регуляция транскрипции.
Негистоновые белки плохо охарактеризованы. Часть негистоновых белков - регуляторы, узнающие определённые нуклеотидные последовательности в ДНК.
СТРУКТУРА ХРОМОСОМ
Фибрилла с нуклеосомными структурами - как бы первичный уровень компактизации ДНК. Другой крайний уровень такой плотной укладки - митотическая хромосома.
Известно, что на одну хромосому эукариотической клетки в среднем приходится несколько сантиметров ДНК (самая большая хромосома дрозофилы содержит около 2 см ДНК). Однако размер митотических хромосом очень мал: несколько мкм в длину при толщине 0,5-1 мкм. Соответственно, ДНК в такой хромосоме уложена с коэффициентом компактизации, равным 10000. Такая плотная упаковка необходима для того чтобы хромосомный материал не разрушался при движении хромосом во время митоза.
Хромосомный цикл

Собственно хромосомы как чёткие, плотные, хорошо видимые в световой микроскоп тела выявляются только незадолго перед клеточным делением. В интерфазе хромосом как плотных тел не видно, т.к. они находятся в разрыхлённом, деконденсированном состоянии. В интерфазе происходит репликация хромосом. Этот период характеризуется синтезом ДНК - синтетический, или S-период. После его окончания кол-во ДНК в интерфазном ядре равно 4c. Собственно хромосомы как нитевидные плотные тела начинают обнаруживаться микроскопически в начале профазы митотического деления. Может сложиться ложное впечатление о том, что кол-во хромосом в профазе равно 2n, но это не так: в профазе каждая хромосома двойная за счёт редупликации. На этой стадии пара хромосом тесно соприкасается друг с другом, взаимно спирализуясь одна относительно другой. Позднее хромосомы в каждой паре начинают обосабливаться, раскручиваться. Становится видно, что кол-во хромосом в начинающей делиться клетке равно 4n. Следовательно, уже в начале профазы хромосомы состояли из двух сестринских хромосом, или хроматид. В профазе и метафазе хромосомы остаются парно связанными друг с другом. В метафазе происходит выстраивание хромосом в экваториальной плоскости клетки и окончательное их разъединение. Клетки остаются в этих фазах тетраплоидными. В анафазе идёт расхождение каждой хромосомы из пары к противоположным полюсам клетки. В телофазе разошедшиеся диплоидные наборы хромосом начинают деконденсироваться. Отдельные хромосомы теряют свои чёткие очертания и теперь уже внутри нового интерфазного диплоидного ядра с 2c ДНК трудно узнать хромосомы, которые можно было видеть во время митоза.
ГЕТЕРОХРОМАТИН И ЭУХРОМАТИН
Термин "гетерохроматин" вначале был предложен для обозначения участков хромосом, интенсивно связывающихся с красителями (т.к. при переходе клетки от митоза к интерфазе определённые зоны различных хромосом или даже целые хромосомы остаются конденсированными - гетерохроматические). Гетерохроматические участки обычно располагаются в теломерных, центромерных, околоядрышковых районах хромосом, но мог






