ГЕНОФОНД БЕЛОРУСОВ
ПО ДАННЫМ О ТРЕХ ТИПАХ ГЕНЕТИЧЕСКИХ МАРКЕРОВ –
АУТОСОМНЫХ, МИТОХОНДРИАЛЬНЫХ, Y ХРОМОСОМЫ
Балановский О.П.1, Тегако О.В. 2
1Государственное учреждение Медико-генетический научный центр РАМН, Москва (Россия)
2Институт искусствоведения, этнографии и фольклора НАН Беларуси, Минск (Беларусь)
ВВЕДЕНИЕ
Изучение этногенеза и этнической истории представляет важную фундаментальную проблему, к решению которой все шире привлекается не только антропология, но и антропогенетика. Среди обширного перечня исторических источников важное место начинают занимать данные о структуре генофонда народа – генетических особенностях населения в различных частях этнического ареала. Генофонд хранит генетическую память об этногенезе. Генетическая летопись, передаваясь в непрерывной цепи поколений, фиксирует события, связанные с перемещениями людей. Это позволяет выделить именно те явления в генофонде, которые неразрывно связаны с демографической историей населения, его миграциями и смешениями, динамикой численности, контактами с соседними этносами и процессами внутренней дифференциации.
Современная антропогенетика стремительно совершенствует инструменты исследования этногенеза, каждый из которых позволяет реконструировать разные аспекты этнических процессов. Наибольшая информативность достигается параллельным анализом всех трех основных типов генетических маркеров, отражающих три главных магистрали микроэволюции популяций:
1) Митохондриальная ДНК – только материнские линии передачи наследственной информации (дочери получают эти гены только от матери, она – от своей матери, и так непрерывная цепь передачи информации по материнской линии приходит из глубины времен).
2) Y хромосома – только отцовские линии передачи наследственной информации (сыну – только от отца, ему – от своего отца, и так непрерывная цепь передачи информации по отцовской линии);
3) Аутосомные маркеры – микроэволюция основной части генофонда (к аутосомным маркерам относятся подавляющая часть генома, переданная от обоих родителей, но рекомбинирующая, то есть «перетасовывающаяся» в процессе передачи информации).
Гаплогруппы Y хромосомы занимают особое место. Уникальная возможность дифференцировать миграции в пространстве и времени, огромная межпопуляционная изменчивость, географические тренды в частотах гаплогрупп – все эти особенности Y хромосомы несут важную информацию о процессах формирования генофонда. Однако маркеры Y хромосомы фиксируют лишь одну своеобразную линию микроэволюции. Чтобы иметь полную и объективную картину, надо оценить микроэволюционные траектории и по другим типам маркеров. Лишь свидетельства всех этих «очевидцев» в комплексе могут дать объективную картину структуры белорусского генофонда и его положения среди других народов Европы. Интенсивными исследованиями, проводимыми ведущими лабораториями мира, уже охвачены практически все народы Европы и смежных регионов. Лишь белорусы являлись одним из немногих народов, до настоящего времени практически неизвестных в отношении полиморфизма этих ДНК маркеров. Заполнение этого «белого пятна» является приоритетным направлением науки.
МАТЕРИАЛЫ
В 2004-2005 годах проведено полевое обследование генофонда белорусов. Сбор образцов венозной крови осуществлен среди коренного сельского населения двух популяций, представляющих основные географические и этнографические подразделения белорусского этноса: северных (Витебская область) и южных (Брестская и Гомельская области) белорусов. Для обследования выбраны районы с высокой долей коренного неметисированного населения. Изучен ряд несмежных населенных пунктов для получения репрезентативной характеристики региона в целом, а не отдельных локальностей. Все обследованные индивидуумы анкетированы по единой программе. В итоговую выборку включены только пробанды, все предки которых на глубину трех поколений происходят из данного региона и являются белорусами. Обследование проведено на основе принципа информированного согласия и под контролем Этической комиссии ГУ МГНЦ РАМН: каждый обследуемый давал письменное информированное согласие на данное обследование. Забор венозной крови осуществлялся системой «Вакуэт» для соблюдения стерильности на уровне европейских стандартов.
Для сравнительного исследования привлечены полностью сопоставимые данные по 14 русским популяциям [Balanovsky et al., 2008] и 5 украинским популяциям [Пшеничнов, 2007], полученными нашим коллективом строго по той же программе, что и для генофонда белорусов.
Генотипирование маркеров Y хромосомы и митохондриальной ДНК проведено О.П. Балановским, аутосомных ДНК маркеров – Мансуровым Р.И., Соловьевой Д.М., Дибировой Х.Д., Кузнецовой М.А., Фроловой С.А. [Мансуров и др., 2006].
ГЕНЕТИЧЕСКИЙ ЛАНДШАФТ Y ХРОМОСОМЫ
Для Европейского субконтинента при изучении генофондов наиболее информативны маркеры Y хромосомы. Распределение гаплогрупп митохондриальной ДНК (мтДНК) и аутосомных маркеров в европейских популяциях, к сожалению, единообразно и требуется прилагать специальные усилия, чтобы выявить различия между популяциями разных народов. Напротив, по Y хромосоме межэтнические различия огромны, и тем самым Y хромосома является идеальной генетической системой для изучения истории генофонда: прослеживания миграций населения, смешения различных групп, действия генетико-автоматических факторов.
В генофонде белорусов нами обнаружены 12 гаплогрупп Y хромосомы: E 3 b 1, E 3 b 3, G 2, I (xI 1 a, I 1 b, I 1 c), I 1 a, I 1 b, I 1 c, J 1, J 2, N 3, R 1 a, R1b3. Наиболее распространены у белорусов четыре гаплогруппы: R 1 a, I 1 b, N 3, R 1 b 3 (частота каждой из этих гаплогрупп – более 5%, гаплогруппы перечислены в порядке убывания частоты). Наибольшая часть генофонда белорусов представлена гаплогруппой R 1 a (60%), и гаплогруппой I 1 b (17%).
Для исследования характера распространения этих гаплогрупп в других частях Евразии были привлечены литературные данные. Высокие частоты гаплогруппы R 1 a типичны как для славяноязычных народов (в том числе и всех восточных славян), так и для некоторых тюркоязычных народов Северной Евразии. Эта гаплогруппа вообще широко распространена в Евразии, но внутри Европы более характерна для восточной, чем для западной её части. Гаплогруппа I 1 b достигает максимальной частоты у хорватов и боснийцев и наиболее распространена в балканском регионе (от 15 до 40%). За пределами этого региона она встречена с меньшей частотой (7-17%) у всех славяноязычных народов Восточной Европы. Гаплогруппа N 3 характерна для финноязычных народов Восточной Европы и балтов: на границе ареалов белорусов и балтов происходит резкое падение частоты N 3. Гаплогруппа R 1 b 3 встречается у белорусов с небольшой частотой: это гаплогруппа, более характерная для населения Западной Европы.
Итак, по спектру гаплогрупп Y хромосомы белорусский генофонд предстаёт как европейский (R 1 b 3), восточноевропейский (по преобладанию R 1 a) и северный восточноевропейский (по присутствию N 3). Гаплогруппа I 1 b отражает связь с миром Южной Европы.
Южные и северные белорусы очень похожи друг на друга: основная часть генофонда маркеров Y хромосомы в обеих изученных белорусских популяциях представлена одним и тем же набором гаплогрупп и близкими частотами. Отличия двух белорусских популяций друг от друга выявляются на уровне тенденций. Несколько большая частота гаплогруппы N3 на севере может быть связана с большей ролью субстратного дославянского населения в формировании современного генофонда. В большей частоте гаплогруппы R1b3 на юго-западе, чем на севере прослеживается общеевропейский тренд изменчивости частоты этой гаплогруппы.
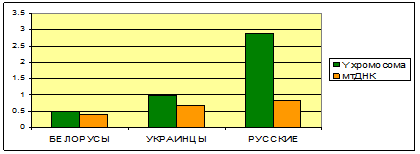
С КЕМ СХОДНЫ БЕЛОРУССКИЕ ПОПУЛЯЦИИ ПО Y ХРОМОСОМЕ?
По результатам многомерного анализа, обе белорусские популяции, будучи географически удалены, тем не менее, чрезвычайно сходны генетически. Несмотря на большое географическое расстояние между изученными белорусскими популяциями, генетическое расстояние между ними меньше среднего расстояния между украинскими популяциями или между русскими популяциями. Генетическое сходство между белорусскими популяциями особенно ярко проявляется на фоне общего европейского разнообразия Y хромосомы.
Различия между северными и юго-западными белорусами всё же проявляются – в большем тяготении северных белорусов к русским Тверской области, а южных белорусов – к русским Воронежской области. Таким образом, сходны популяции, находящиеся примерно на одной географической широте, в то время как между популяциями разных широт наблюдаются большие различия.
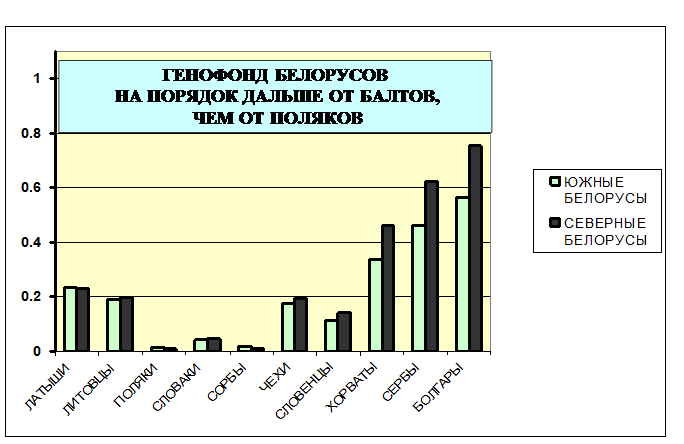
Рис. 1. Генетические расстояния от белорусских популяций до народов Европы по данным о гаплогруппах Y хромосомы (отцовская линия наследования)
Светлые столбцы - генетические расстояния от южных белорусов.
Темные столбцы - генетические расстояния от северных белорусов.
Сравним теперь генофонд белорусов с другими генофондами народов Европы (рис. 1). Поскольку предполагается, что балтский субстрат может быть существенен в генофонде белорусов, рассмотрим – насколько близок генофонд белорусов к современным балтам? Мы видим, что генетические расстояния до балтов – и от северных, и от южных белорусов – одинаково велики: до латышей d=0.23, до литовцев d=0.20. До многих популяций западных славян это расстояние на порядок меньше: до поляков d=0.01, до сорбов (лужичан) d=0.02, словаков d=0.04. От белорусов до украинцев в среднем генетическое расстояние столь же мало: d=0.04, варьируя между разными популяциями от d=0.01 (между южными белорусами и подольскими украинцами) до d=0.08 (между северными белорусами и западными украинцами).
От белорусов до русских популяций в среднем генетическое расстояние немногим больше: d=0.06. Но если исключить генетически резко своеобразные северные русские популяции (расстояние от белорусов до них составляет около d=0.17), то расстояние до остальных русских популяций снизится всего лишь до d=0.02.
Таким образом, по наиболее генетически информативным маркерам Y хромосомы белорусы обнаруживают высокое сходство с восточными славянами и большинством западных славян, но генетически далеки от балтов. Причем удаленность от балтов одинаково велика и для северных, и для южных белорусов.
Белорусские популяции соответствуют основному Евразийскому тренду. В целом в Белоруссии обнаруживается разнообразие по гену устойчивости к СПИДу (CCR 5 del 32) и общеевропейский тренд – падения частоты гена с северо-запада на юго-восток.
Однако наиболее мощным и точным инструментарием геногеографии являются не методы многомерной статистики, а компьютерный картографический анализ. Он проведен для всех типов маркеров и для разных генофондов – белорусов, русских, украинцев, Восточной Европы, Европы в целом и Евразии.
Не имея возможности в статье привести все множество карт (продемонстрированных в докладе и частично приведенных в монографии [Балановская, Балановский, 2007]), укажем основные выводы картографического анализа.
1). Карты главных компонент белорусского генофонда и по данным о классических (иммуно-биохимических) маркерах, и по данным об аутосомных ДНК маркерах выявляют один и тот же главный сценарий изменчивости генофонда белорусов. Четкая широтная изменчивость является главной закономерностью белорусского генофонда: одни частоты генов оказываются характерны для юга Белоруссии, тогда как северная часть ареала занята несколько отличным генофондом. При этом большая часть ареала занята промежуточными значениями, переходными между крайним северным и крайним южным вариантами.
Итак, главный вывод картографического анализа генофонда белорусов: широтная изменчивость – постепенный переход, различия между северными и южными популяциями – является основной закономерностью белорусского генофонда.
2). Другая важная закономерность, выявляемая картами главных компонент, - ядра своеобразных значений расположены не внутри ареала белорусов, а на границах. Они свидетельствуют об интенсивных потоках генов из других этносов и о том, что особенности генофонда белорусов складывались на основе взаимодействия с соседними этносами.
3). Выявленная у белорусов основная закономерность «север - юг» является частью общего восточноевропейского тренда. Хотя в целом он имеет долготную изменчивость «юго-запад - восток» на всем пространстве Восточной Европы, но именно в той его части, где расположен ареал белорусов и их ближайших соседей, он имеет локальные широтные особенности.
4). Насколько такая картина характерна для других восточных славян? Детальный анализ русского генофонда [Балановская, Балановский, 2007] показал, для русских популяций также характерна широтная изменчивость «север-юг». Однако она является своей, собственной, характерной именно для русского генофонда: при анализе всех популяций Восточной Европы в русском ареале обнаруживается не широтный, а долготный восточно-европейский тренд «юго-запад - восток». Предполагается, что такая особенность русского генофонда связана с историей его формирования: двух потоков древних славянских племен – северного (новгородского) и южного. Вторая важная закономерность русского генофонда - ядра своеобразных значений расположены внутри русского ареала, а не на его границах. Это указывает, что не миграционные процессы, а особенности собственного саморазвития русского генофонда сформировали основной генетический ландшафт русского генофонда.
5).Итак, карты главных компонент выявляют сходство главного тренда белорусского генофонда и с русским генофондом, и со всем генофондом Восточной Европы. Это позволяет считать, что белорусский генофонд входит в генетическое пространство и русского генофонда, и генофонда Восточной Европы, а своеобразие на границах ареала белорусов обусловлено контактами с другими этносами (потоки генов).
6). Этот вывод подтвержден и другим независимым анализом – картографированием генетических расстояний. Карты расстояний от русского генофонда ярко демонстрируют, что белорусы входят в генетическое пространство русского генофонда, близкого к генофонду всех восточных славян. Причем белорусские популяции расположены ближе к средним русским показателям, чем многие русские популяции (особенно на севере и востоке русского ареала). В целом, русский генофонд наиболее полно представляет генофонд всех восточных славян, а белорусский генофонд – малую часть его изменчивости.
7). Карты генетических расстояний от белорусского генофонда подтверждают, что белорусы отражают лишь одну из частей генофонда восточных славян и связаны с ними мощными генными потоками. По сравнению с русским генофондом изменчивость белорусского генофонда мала. Обнаруживается широкая переходная зона - плавное «перетекание» белорусского генофонда в генофонды восточных славян.
ЗАКЛЮЧЕНИЕ.
Полученные нами новые данные о белорусских популяциях позволили заполнить «белое пятно» на генетической карте Восточной Европы. Благодаря этому, было выявлено существование генетической общности, занимающей обширную территорию.
По маркерам Y хромосомы при сравнении белорусских популяций с популяциями Восточной, Западной, Северной и Южной Европы, выявлена генетическая общность, охватывающая территорию от Польши до запада Центральной России, включая Белоруссию. На юге граница общности проходит между ареалом белорусов и украинцев (возможно, область колебаний границы леса и лесостепи в исторический период), на северо-востоке – между белорусами и северо-западной областью русского ареала. Общность образует единый плотный кластер, включающий белорусов, поляков, русских юго-западных областей (Белгородской, Воронежской, Курской, Орловской областей), а также русских Тверской области.
По частотам гаплогрупп мтДНК выявлены различия между южными и северными белорусами. Они формируются за счет выраженного своеобразия северных белорусов.
По гену устойчивости к СПИДу (CCR5del32) генофонд белорусов следует Евразийскому тренду. Между белорусскими популяциями наблюдаются различия, соответствующие общеевропейскому тренду: падение частоты гена с северо-запада на юго-восток.
По большой панели аутосомных ДНК маркеров все восточные славяне входят в один кластер Восточной Европы.
По большой панели классических маркеров, как и по аутосомным ДНК маркерам, все восточные славяне входят в один кластер. При этом генетические различия между белорусскими популяциями невелики: и южные, и северные белорусы входят в единый кластер.
Картографический анализ генофонда белорусов показал, что основной закономерностью белорусского генофонда является широтная изменчивость – постепенный переход между северными и южными популяциями.
Сравнительный картографический анализ с другими генофондами Европы показал, что белорусский генофонд входит в генетическое пространство и русского генофонда, и генофонда Восточной Европы. При этом генофонд белорусов объемлет небольшую часть изменчивости общего восточнославянского генофонда.
Сформулируем краткие ответы на некоторые вопросы истории формирования генофонда белорусов.
Литература
Балановская Е.В., Балановский О.П. Русский генофонд на Русской равнине. М.: Луч, 2007. 416 с.
Балановский О.П. База данных о полиморфизме митохондриальной ДНК в населении мира – новый Интернет-ресурс для исследований в популяционной, медицинской генетике и криминалистике // Медицинская генетика. 2005. № 4. С. 154.
Балановский О.П., Роотси С., Чурносов М.И., Почешхова Э.А., Болдырева М.Н., Евсеева И.В., Виллемс Р., Балановская Е.В. К вопросу о таксономической ценности разных классов ДНК маркеров: закономерная географическая изменчивость Y хромосомы // VII Конгресс этнографов и антропологов России. Саранск. 2007. С. 267.
Мансуров Р.И., Соловьёва Д.С., Тегако О.В., Дибирова Х.Д., Пшеничнов А.С., Кузнецова М.А., Балановский О.П., Воронько О.Е., Фролова С.А., Балановская Е.В. Генофонд северных и южных белорусов по аутосомным ДНК маркерам (7 локусов инсерционно-делеционного полиморфизма) // Материалы Международной Конференции "Генетика в России и мире". Москва, 2006.
Пшеничнов А.С. Структура генофонда украинцев по данным о полиморфизме митохондриальной ДНК и Y хромосомы. Автореф. дис. канд. биол. наук. / ГУ Медико-генетический научный центр РАМН. М., 2007. 29 с.
Balanovsky O., Pocheshkhova E., Pshenichnov A., Solovieva D., Kuznetsova M., Voronko O., Churnosov M., Tegako O., Atramentova L., Lavryashina M., Evseeva I., Borinska S., Boldyreva M., Dubova N., Balanovska E. Is spatial distribution of the HIV-1-resistant CCR5Delta32 allele formed by ecological factors? // J Physiol Anthropol Appl Human Sci. 2005 Jul. V 24. N 4. P. 375-382.
Balanovsky O., Pshenichnov A., Rootsi S., Churnosov M., Atramentova L., Tegako L., Yankovsky N. Intra-ethnic variation of the Y chromosome in European countries: a comparative study // ISABS, Split. 2007. International Society for Applied Biological Sciences, Split. 2007. P. 121.
Balanovsky O., Rootsi S., Pshenichnov A., Kivisild T., Churnosov M., Evseeva I., Pocheshkhova E., Boldyreva M., Yankovsky N., Balanovska E., Villems R.. Two sources of the Russian patrilineal heritage in their Eurasian context // Am J Hum Genet. 2008, V. 82. № 1. P.236-250.
Galvani A.P., Novembre J. The evolutionary history of the CCR5-Delta32 HIV-resistance mutation // Microbes Infect. 2005. N 7(2). Р. 302-309.
Libert F., Cochaux P., Beckman G., Samson M., Aksenova M., Cao A., Czeizel A., Claustres M., de la Rúa C., Ferrari M., Ferrec C., Glover G., Grinde B., Güran S., Kucinskas V., Lavinha J., Mercier B., Ogur G., Peltonen L., Rosatelli C., Schwartz M., Spitsyn V., Timar L., Beckman L., Parmentier M., Vassart G. The deltaccr5 mutation conferring protection against HIV-1 in Caucasian populations has a single and recent origin in Northeastern Europe // Hum. Mol. Genet. 1998. N 7 (3). P. 399-406.
Martinson J.J., Chapman N.H., Rees D.C., Liu Y-T., Clegg J.B. Global distribution of the CCR5 gene 32-basepair deletion // Nature Genet. 1997. V.16. P. 100-103.
Martinson J.J., Hong L., Karanicolas R., Moore J.P., Kostrikis L.G. Global distribution of the CCR2-64I/CCR5-59653T HIV-1 disease-protective haplotype // AIDS. 2000. N 14(5). P. 483-489.
Stephens J.C., Reich D.E., Goldstein D.B., Shin H.D., Smith M.W., Carrington M., Winkler C., Huttley G.A., Allikmets R., Schriml L., Gerrard B., Malasky M., Ramos M.D., Morlot S., Tzetis M., Oddoux C., di Giovine F.S., Nasioulas G., Chandler D., Aseev M., Hanson M., Kalaydjieva L., Glavac D., Gasparini P., Kanavakis E., Claustres M., Kambouris M., Ostrer H., Duff G., Baranov V., Sibul H., Metspalu A., Goldman D., Martin N., Duffy D., Schmidtke J., Estivill X., O'Brien S.J., Dean M. Dating the origin of the CCR5-32 AIDS-resistance allele by the coalescence of haplotypes // Am. J. Hum. Genet. 1998. V. 62. P. 1507-1515
ГЕНОФОНД БЕЛОРУСОВ
ПО ДАННЫМ О ТРЕХ ТИПАХ ГЕНЕТИЧЕСКИХ МАРКЕРОВ –







