

Своеобразие русской архитектуры: Основной материал – дерево – быстрота постройки, но недолговечность и необходимость деления...

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Своеобразие русской архитектуры: Основной материал – дерево – быстрота постройки, но недолговечность и необходимость деления...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Топ:
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Когда производится ограждение поезда, остановившегося на перегоне: Во всех случаях немедленно должно быть ограждено место препятствия для движения поездов на смежном пути двухпутного...
Интересное:
Влияние предпринимательской среды на эффективное функционирование предприятия: Предпринимательская среда – это совокупность внешних и внутренних факторов, оказывающих влияние на функционирование фирмы...
Лечение прогрессирующих форм рака: Одним из наиболее важных достижений экспериментальной химиотерапии опухолей, начатой в 60-х и реализованной в 70-х годах, является...
Аура как энергетическое поле: многослойную ауру человека можно представить себе подобным...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
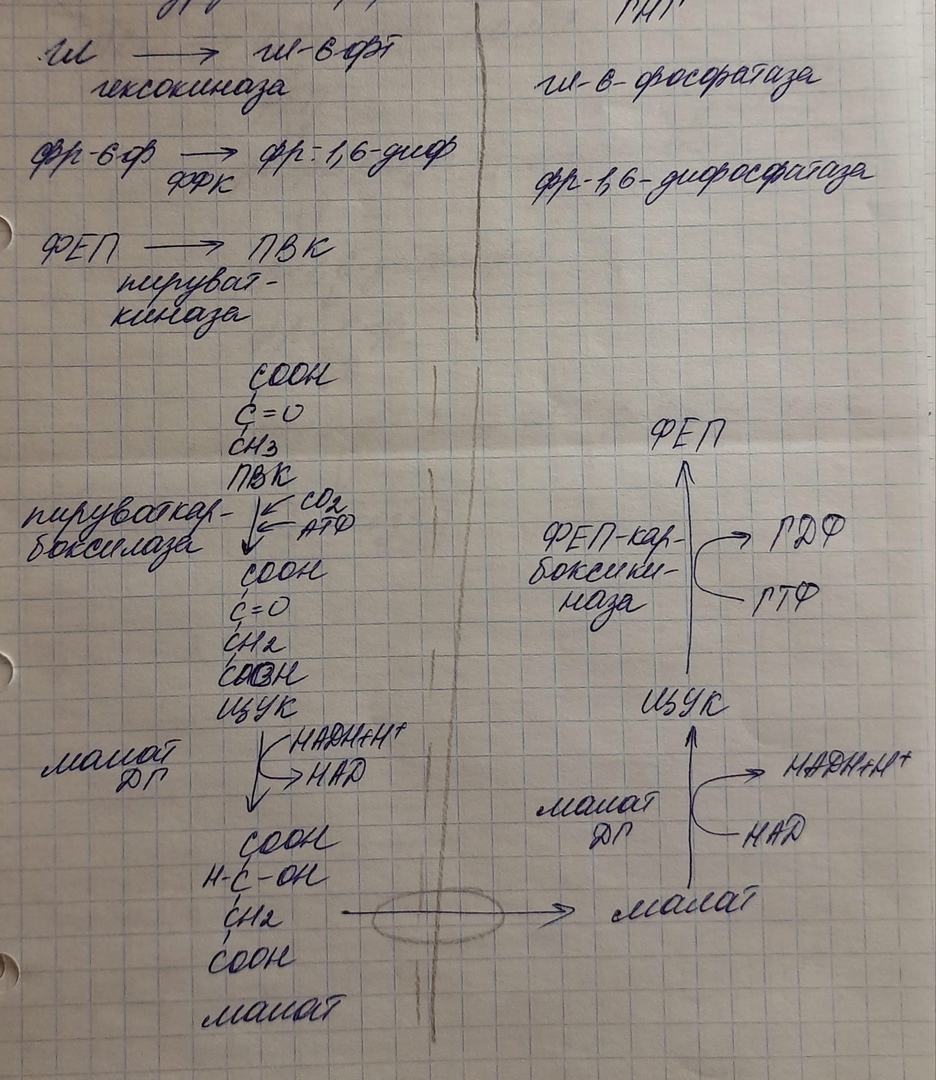
Последняя реакция катализируется глюкозо-6-фосфатазой. Она имеется только в печени и почках, следовательно, только эти ткани могут продуцировать свободную глюкозу. 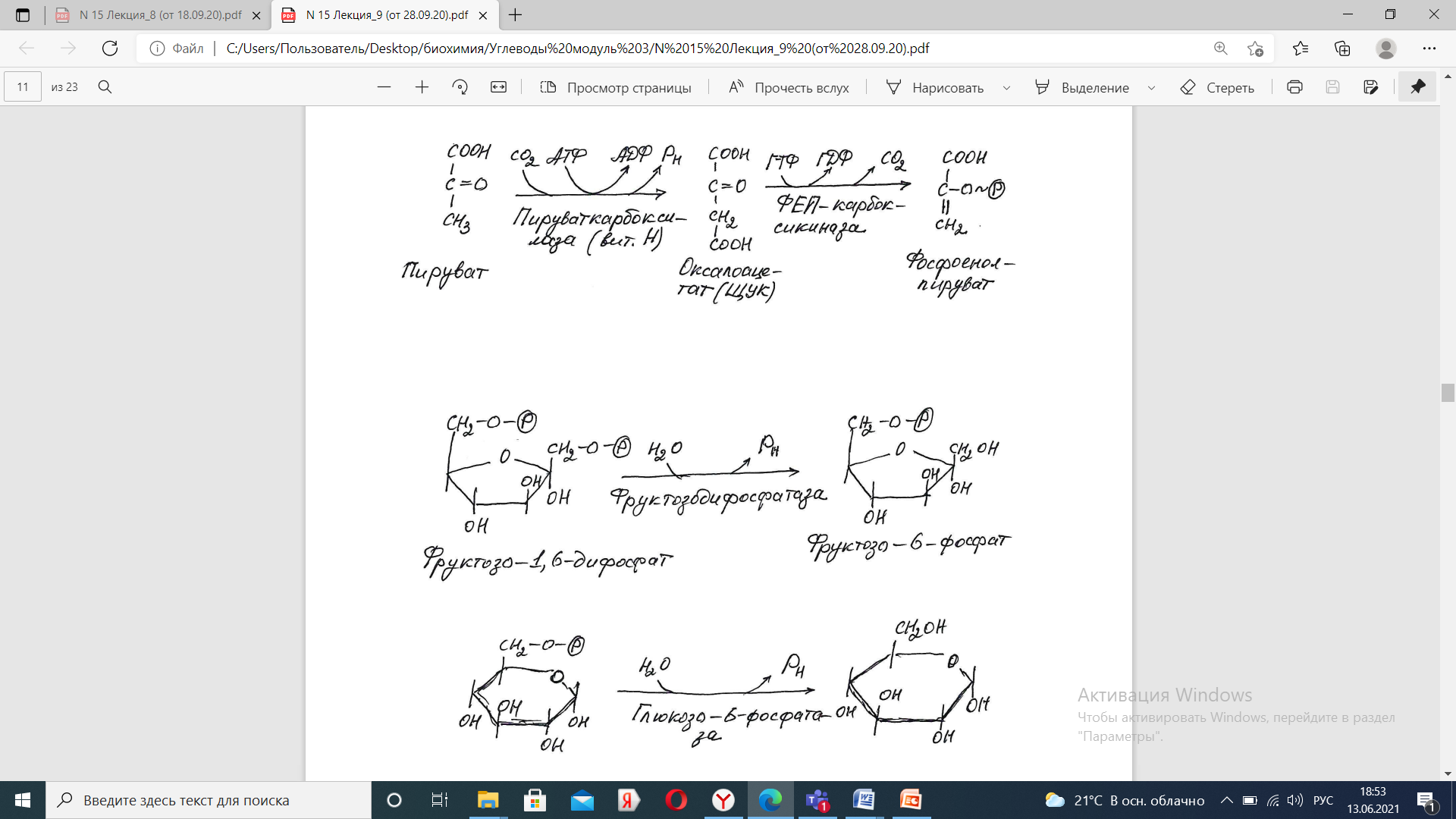
Регуляция ГНГ
1. ↓АТФ и НАД+ - тормозится.
2. Ключевые ферменты ГНГ:
- активируют: АТФ, глюкокортикоиды, ЖК, глюкагон;
- ингибируют: АДФ и АМФ, инсулин.
Кроме гормонов, имеются метаболическая регуляция, при которой аллостерически активируется пируваткарбоксилаза при помощи ацетил-SКоА, фруктозо-1,6-дифосфатаза при участии АТФ.
В качестве примера синтеза глюкозы из аминокислот рассмотрим участие в этом процессе глутамата, аспартата, серина и аланина.
В реакциях глюконеогенеза разрывается 6 макроэргических фосфатных связей связей в 4 молекулах АТФ и 2 молекулах ГТФ. Для синтеза 1 молекулы глюкозы необходимы 2 молекулы пирувата.
Запишем суммарное уравнение глюконеогенеза:
2 Пируват + 2NADH + 4H+ + 4АТФ + 2ГТФ + 6H2O → Глюкоза + 2NAD+ + 4АДФ + 2ГДФ + 6Фн
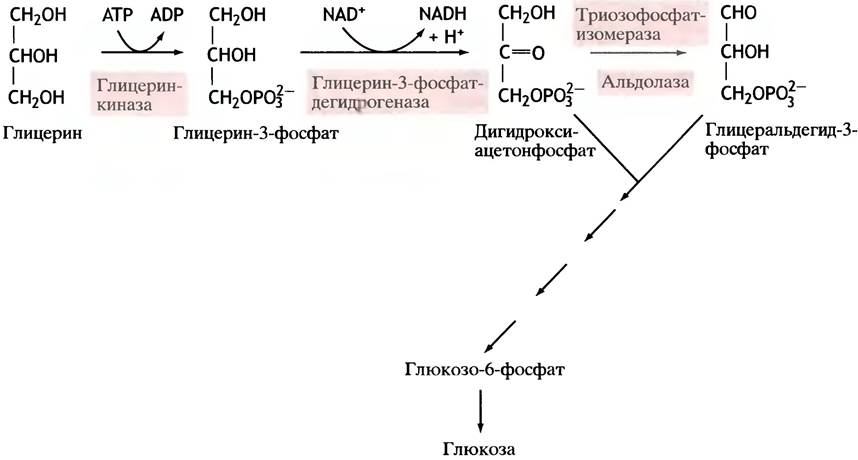
На карбоксилирование 2 молекул пирувата затрачивается 2 молекулы АТФ. Далее 2 молекулы ГТФ требуются для образования фосфоенолпирувата из оксалоацетата. И ещё 2 молекулы АТФ необходимы для фосфорилирования 2 молекул 3-фосфо-глицерата до 1,3-бисфосфоглицерата. Энергия восстановительных эквивалентов (NADH) нужна для образования глицеральдегид-3-фосфата.
При голодании глюконеогенез обеспечивается энергией с помощью β-окисления жирных кислот.
Если привести все энергоносители к одному знаменателю (АТФ), то в ходе ГНГ затрачивается:
1. 4 молекулы АТФ и 2 молекулы ГТФ (эквивалентны 2 молекулам АТФ);
2. 2 молекулы NADH (эквивалентны 6 молекулам АТФ).
Суммируем все затраты и получаем: 4 АТФ + 2 АТФ + 6 АТФ = 12 АТФ
Итак, в ходе глюконеогенеза затрачивается 12 молекул АТФ.
|
|
54.Апотомическое окисление глюкозы (пентозный цикл). Окислительная и неокислительная фазы, химизм и ферменты парциальных реакций. Роль пентозофосфатного пути, связь с процессами синтеза нуклеотидов, жирных кислот, дихотомического окисления глюкозы, микросомального окисления.
АПОТОМИЧЕСКИЙ ПУТЬ ОКИСЛЕНИЯ ГЛЮКОЗЫ (Пентозофосфатный путь, ПФП)
Протекает в цитозоле клеток печени, жировой ткани, эритроцитах, коре надпочечников, молочной железе при лактации, в скелетных мышцах.
Биологическое значение:Роль:
1. Образуется НАДФН: используются на синтеза ЖК, ХС, желчных кислот, стероидных гормонов, АК, обезвреживании токсических соединений.
2. Образуется рибозо-5-фосфат: используется на синтез нуклеотидов, НК, кофакторов.
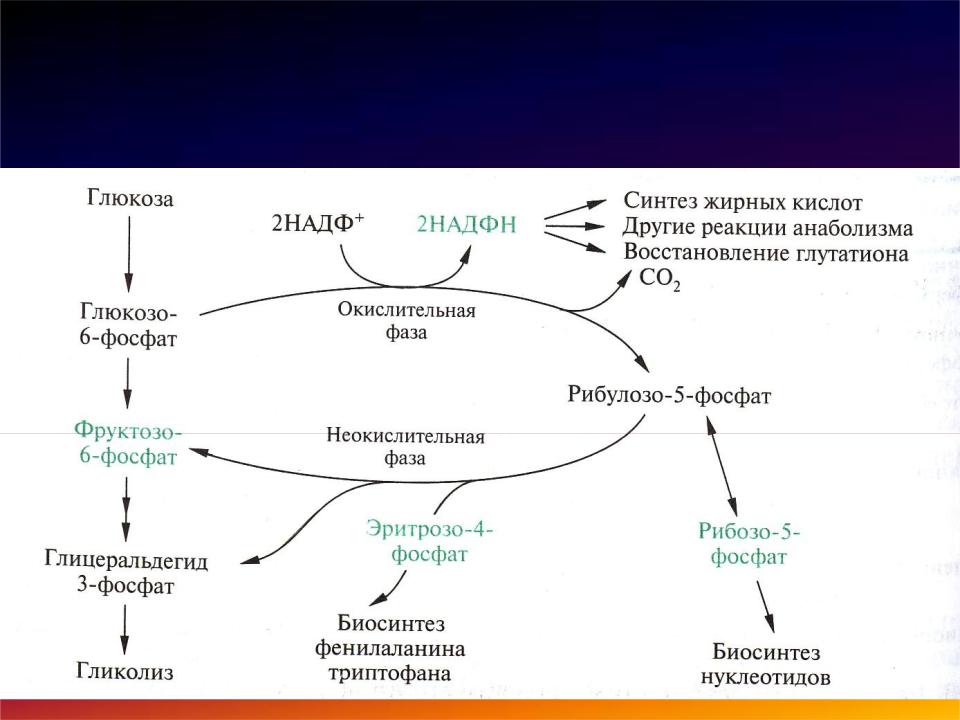
ПФП включает два этапа – окислительный и неокислительный.
Суммарное уравнение ПФП: 6 глюкозы + 12 НАДФ → 5 глюкозы + 6 СО2 + 12 НАДФН2
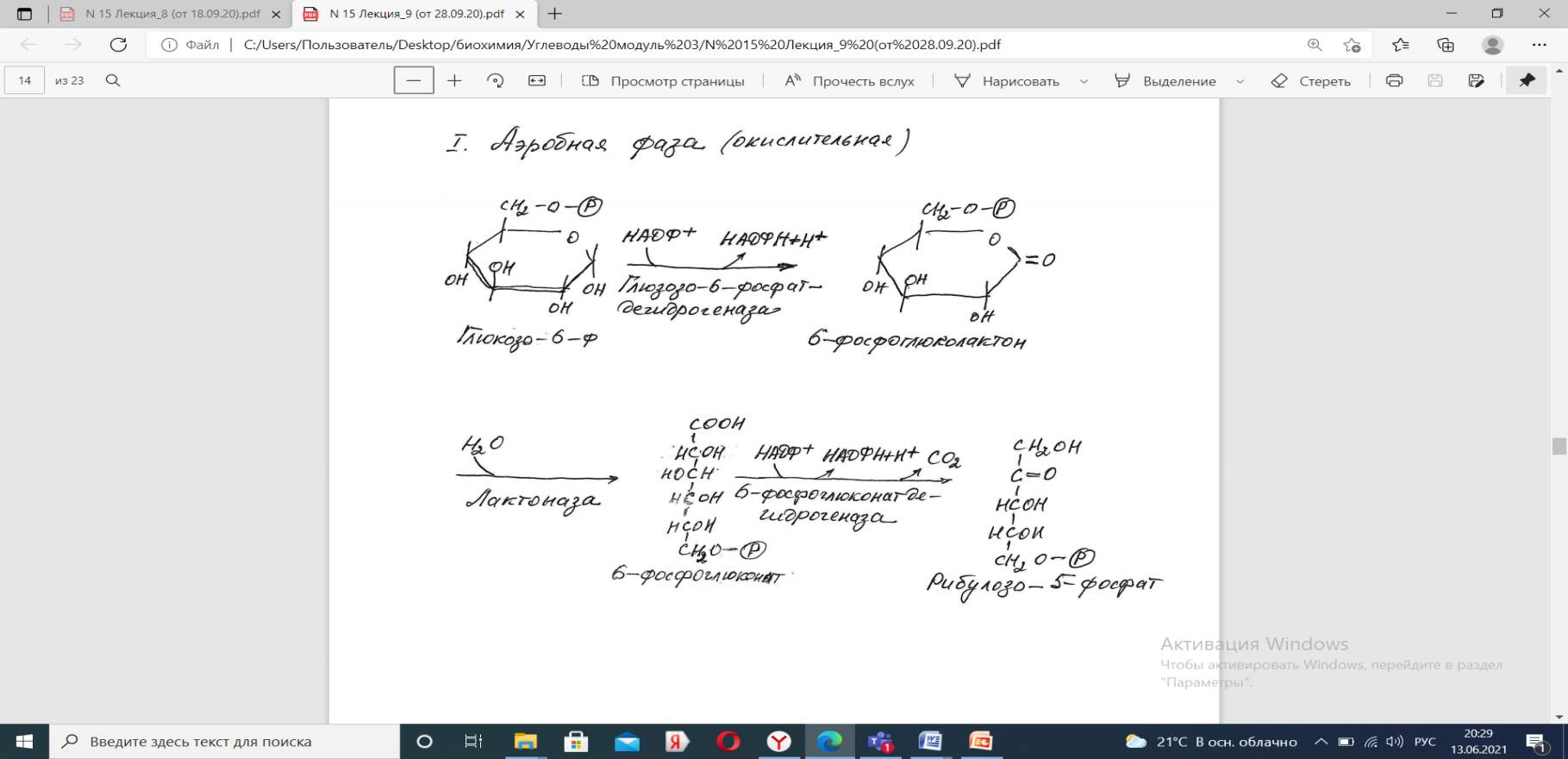
На первом, окислительном, этапе глюкозо-6-фосфат в трех реакциях превращается в рибулозо-5-фосфат, реакции сопровождаются восстановлением двух молекул НАДФ до НАДФН.
На этом этапе происходит регуляция процесса: инсулин повышает активность глюкозо-6-фосфат-дегидрогеназы и фосфоглюконат-дегидрогеназы.
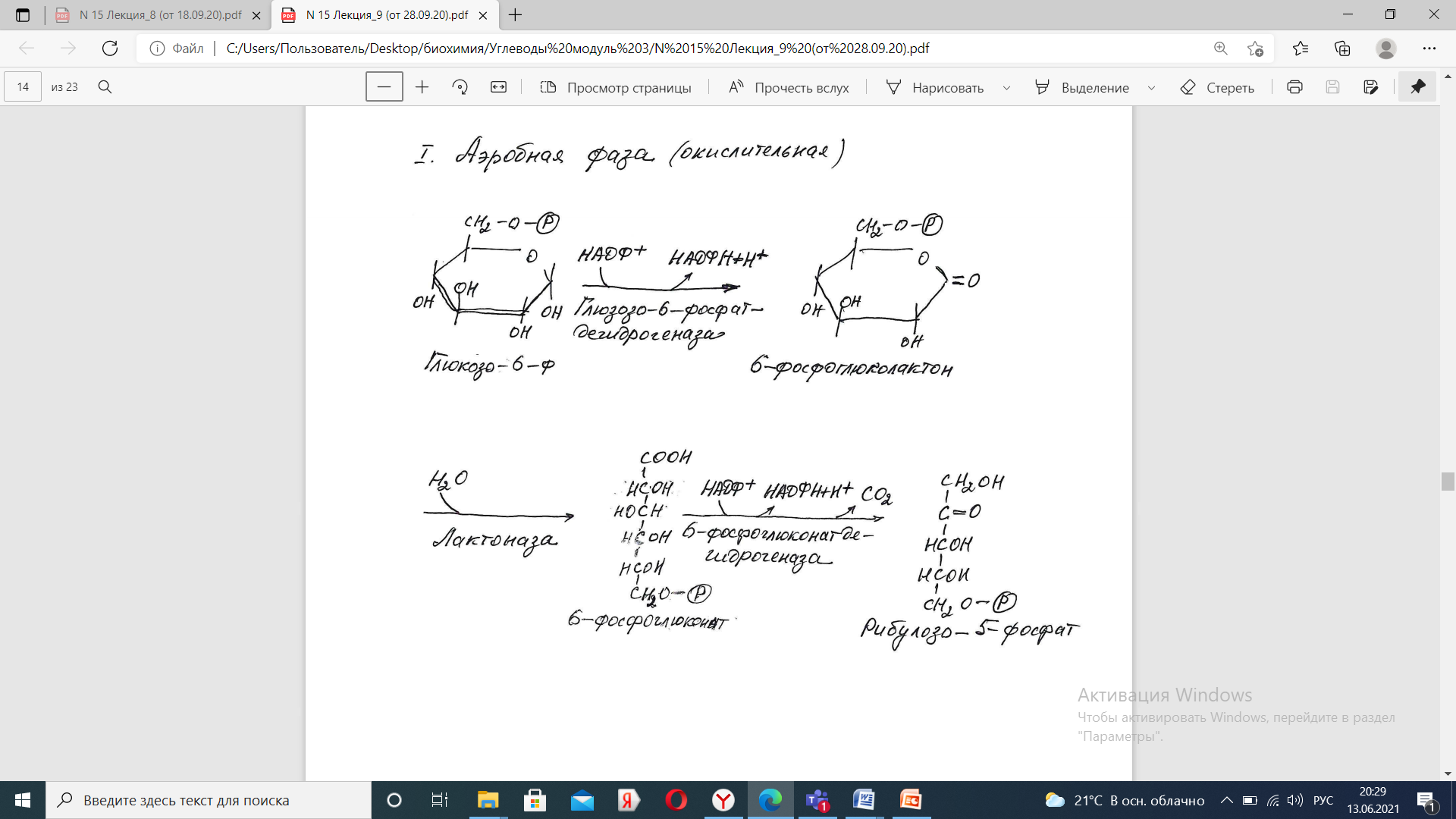
Неокислительная фаза- Второй этап – этап структурных перестроек, благодаря которым пентозы способны возвращаться в фонд гексоз. В этих реакциях рибулозо-5-фосфат изомеризуется до рибозо- 5-фосфата и ксилулозо-5-фосфата. Далее под влиянием ферментов транскетолазы и трансальдолазы происходят структурные перестройки с образованием других моносахаридов.
При реализации всех реакций второго этапа пентозы превращаются во фруктозо-6-фосфат и глицеральдегидфосфат. Из глицеральдегид-3-фосфата при необходимости могут образоваться гексозы.

Ключевые ферменты:
1) глюкозо-6-фосфатдегидрогеназа — главный ключевой фермент;
2) 6-фосфоглюконатдегидрогеназа.
Значение окислительного этапа:
1. Главный поставщик рибозо-5-фосфата для биосинтетических процессов:
|
|
o биосинтез мононуклеотидов (АМФ, ГМФ, УМФ, ЦМФ, ТМФ и др.);
o синтез нуклеиновых кислот (ДНК, РНК);
o синтез коферментов (НАД+, НАДФ+, ФАД, КоА-SН).
2. Основной источник НАДФН.Н+ в клетках. ПФП на 50 % обеспечивает потребности клетки в НАДФН.Н+.
НАДФН.Н+ в клетках используется:
1) в реакциях биосинтеза веществ как восстановитель:
•
o синтез жирных кислот;
o биосинтез холестерола, стероидных гормонов, желчных кислот;
o синтез заменимых аминокислот (НАДФН•Н+ как кофермент глутаматдегидрогеназы в реакциях восстановительного аминированияα-кетоглутаровой кислоты);
o в глюкуроновом пути и др.
2) в обезвреживании веществ: в реакциях гидроксилирования различных ксенобиотиков, лекарственных веществ, этанола и других веществ, которые осуществляются с участием микросомной цитР450-зависимой системы окисления;
3) как антиоксидант: используется на восстановление окисленногоглутатиона. Глутатион — важного антиоксиданта клеток;
4) в фагоцитозе: генерирование активных форм кислорода. Фагоциты с использованием НАДФН.Н+ генерируют супероксидные анион-радикалы, выполняющие основную роль в разрушении поглощённых бактериальных клеток. При недостаточной продукции НАДФН.Н+ при нарушении ПФП отмечается хроническое течение инфекционных заболеваний.
Интенсивность протекания реакций ПФП зависит от потребности клеток в продуктах реакций и различается в разных тканях. Реакции окислительного этапа активно протекают в клетках печени, жировой ткани, эмбриональной ткани, в коре надпочечников, щитовидной железе, половых железах, лактирующей молочной железе, костном мозге, эритроцитах.
II. Неокислительный этап (этап межмолекулярных перегруппировок). На этом этапе происходят взаимопревращения сахаров (фосфотриоз, фосфотетроз, фосфопентоз, фосфогексоз, фосфогептулоз, фосфооктулоз), в результате которых регенерирует глюкозо-6-фосфат.
Два основных фермента катализируют превращения на неокислительном этапе:
1) транскетолаза катализирует перенос двухуглеродных фрагментов. В качестве кофермента использует тиаминпирофосфат;
2) трансальдолаза катализирует перенос трёхуглеродных фрагментов.
Варианты неокислительных превращений:
• классический или F-вариант (от англ. fat — жир) — осуществляется в клетках жировой ткани;
|
|
• октулозный или L-вариант (от англ. liver — печень) — осуществляется в клетках печени и других тканей.
Итак, на неокислительном этапе невостребованные в клетках пентозофосфаты в результате межмолекулярных перегруппировок превращаются в Гл-6-Ф, а также образуются Фр-6-Ф и 3-ФГА.
Все реакции неокислительного этапа обратимы.
На неокислительном этапе ПФП связан с гликолизом (посредством Гл-6-Ф, Фр-6-Ф и 3-ФГА), то есть возможно переключение этих процессов.
Значение неокислительного этапа:
1. Стабилизирует концентрацию фосфопентоз в клетке, то есть утилизирует лишниефосфопентозы. Благодаря связи с гликолизом лишние пентозы катаболизируют по гликолитическому пути, давая клеткам энергию.
2. Синтез фосфопентоз в клетке при торможении окислительного этапа благодаря обратимости реакций неокислительного превращения.
Регуляция пентозофосфатного пути, в основном, осуществляется на уровне дегидрогеназ. Инсулин индуцирует синтез глюкозо-6-фосфатдегидрогеназы, 6-фосфоглюконат-дегидрогеназы. Жирные кислоты — аллостерические ингибиторы глюкозо-6-фосфат-дегидрогеназы. Увеличение уровня НАДФН.Н+ в клетке тормозит окисление глюкозы по ПФП.
Взаимосвязь дихотомического и апотамического путей окисления глюкозы

| 
|
Регуляция ПФП
Ключевые ферменты: - Глюкозо-6-ф.ДГ,
- 6-ф.глюконатДГ,
- транскетолаза.
Активность ПФП увеличивается:
1) при ↑НАДФ+/НАДФН,
2) под влиянием инсулина и Т3 и Т4 – активаторы;
ГК – ингибиторы.
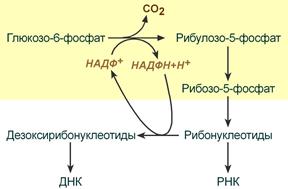
| Растущие и делящиеся клетки Если клетка растет и делится, то ей нужны НАДФН и рибозо-5-фосфат. В этом случае 2-й этап идти не будет, весь образуемый на 1-м этапе рибулозо-5-фосфат превратится в рибозо-5-фосфат. Который далее фосфорилируется в фосфо-рибозил-дифосфат и используется для синтеза как пуриновых, так и пиримидиновых нуклеотидов. НАДФН будет расходоваться на синтез дезоксирибонуклеотидов. При распаде нуклеотидов образуемый рибозо-5-фосфат через второй этап способен превратиться в фр.-6-фосфат и окисляться с получением энергии. | Особенность пентозофосфатного шунта при активном синтезе РНК и ДНК
|
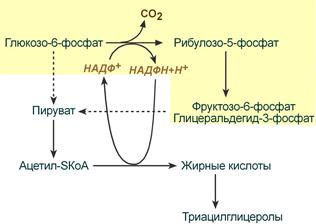
| Адипоциты Если потребность в НАДФН значительно превышает потребность в рибозо-5-фосфате при синтезе жирных кислот, то по окислительным реакциям ПФП образуются НАДФН и рибулозо-5-фосфат. Далее, под действием ферментов 2 этапа, рибулозо-5-фосфат превращается в другие пентозо-5-фосфаты и далее в метаболиты гликолиза (фр.-6-фосфат и глицеральдегид-3-фосфат). Эти метаболиты превращаются в ПВК и ацетил-SKoA и, в основном, используются для синтеза жир.кислот и ХС. Одновременно глицеральдегид-3-фосфат может превращаться в глицерол-3-фосфат и уходить в биосинтез ТАГ. | Особенность ПФП в адипоците
|
| Эритроциты В случае, когда потребность в НАДФН высока, а потребность в рибозо-5-фосфате отсутствует, как в эритроците, где НАДФН активно используется для восстановления антиоксиданта глутатиона ферментом глутатионредуктазой, последовательно идут оба этапа ПФП. Глюкозо-6-фосфат превращается в рибозо-5-Ф и далее во фруктозо-6-фосфат и глицеральдегид-3-фосфат, которые "проваливаются" в гликолиз с образованием лактата. В конечном результате происходит одновременное генерирование НАДФН и АТФ. | Особенность пентозофосфатного шунта в эритроците

|
|
|
55. Особенности обмена фруктозы и галактозы. Врожденные нарушения их обмена. Транзиторные и наследственные фруктозурии, галактоземия и галактозурия. Коррекция этих нарушений. Биологическое значение галактозы и фруктозы в обмене новорожденных.
ПРЕВРАЩЕНИЕ ФРУКТОЗЫ
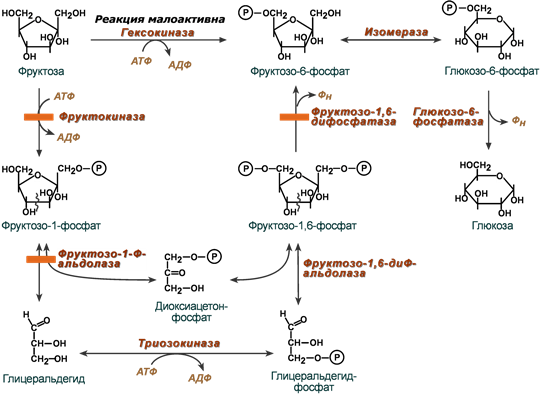

Реакции перехода фруктозы в глюкозу достаточно просты. Сначала происходит активация фруктозы посредством фосфорилирования 6- го атома углерода. В печени реакцию катализируют ферменты – гексокиназа и фруктокиназа, но гексокиназа имеет гораздо более низкое сродство к фруктозе.
Фруктозо-6-фосфат далее изомеризуется и глюкозо -6- фосфатаза отщепляет уже ненужный фосфат.
Особенностью метаболизма фруктозы является то, что фермент фруктокиназа является инсулин - независимым. В результате превращение фруктозы в пировиноградную кислоту и ацетил-SКоА происходит быстрее, чем для глюкозы. Это объясняется "игнорированием" лимитирующей реакции метаболизма глюкозы, катализируемой фосфофруктокиназой.
Дальнейший метаболизм ацетил-SКоА в данном случае может привести к избыточному образованию жирных кислот и триацилглицеролов.
НАРУШЕНИЯ ПРЕВРАЩЕНИЯ ФРУКТОЗЫ:

1.Эссенциальная фруктозурия
Генетический дефект фруктокиназы приводит к доброкачественной эссенциальной фруктозурии, протекающей безо всяких отрицательных симптомов.
2.Наследственная фруктозурия
Заболевание формируется вследствие наследственных аутосомно-рецессивных дефектов других ферментов обмена фруктозы.
Дефект фруктозо -1- фосфатальдолазы проявляется после введения в рацион младенца соков и фруктов, содержащих фруктозу.
|
|
Патогенез связан со снижением концентрации фосфора в крови, гиперфруктоземией, с тяжелой постпрандиальной гипогликемией. Отмечается вялость, нарушения сознания, почечный канальцевый ацидоз. Диагноз ставится исходя из "непонятного" заболевания печени, гипофосфатемии,гиперурикемии, гипогликемии и фруктозурии. Для подтверждения проводят тест толерантности к фруктозе. Лечение включает диету с ограничением сладостей, фруктов, овощей. Дефект фруктозо -1,6- дифосфатазы проявляется сходно с предыдущим, но не так тяжело.
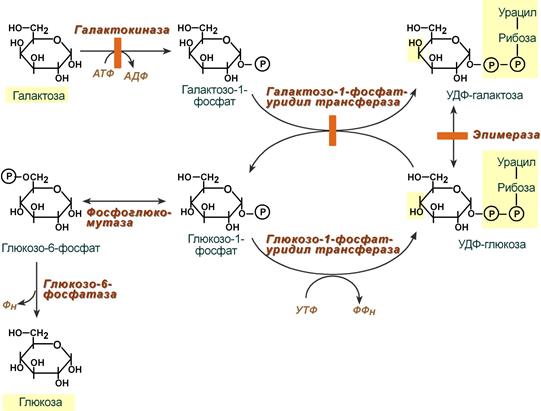
ПРЕВРАЩЕНИЕ ГАЛАКТОЗЫ
| Галактоза сначала подвергается фосфорилированию по 1- му атому углерода. Отличием от обмена фруктозы является превращение в глюкозу не напрямую, а через синтез УДФ-галактозы, которая впоследствии изомеризуется в глюкозу | 
|
|
|
|

Таксономические единицы (категории) растений: Каждая система классификации состоит из определённых соподчиненных друг другу...

Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...

Типы сооружений для обработки осадков: Септиками называются сооружения, в которых одновременно происходят осветление сточной жидкости...

Архитектура электронного правительства: Единая архитектура – это методологический подход при создании системы управления государства, который строится...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!