Систематика микроорганизмов (прокариоты, грибы, простейшие, вирусы). Принципы классификации. Таксономические категории. Понятие о виде как основной таксономической единице. Номенклатура. Основные отличия прокариотов от эукариотов.

Микроорганизмы представлены доклеточными формами (вирусы — царство Vira) и клеточными формами (бактерии, архебактерии, грибы и простейшие). Различают 3 домена (или «империи»): «Bacteria», «Archaea» и «Eukarya»:
Ø □ домен «Bacteria» — прокариоты, представленные настоящими бактериями (эубактериями);
Ø □ домен «Archaea» — прокариоты, представленные архебактериями;
Ø □ домен «Eukarya» — эукариоты, клетки которых имеют ядро с ядерной оболочкой и ядрышком, а
цитоплазма состоит из высокоорганизованных органелл — митохондрий, аппарата Гольджи и др. Домен «Eukarya» включает: царство Fungi (грибы); царство животных Animalia (включает прстейшие – подцарство Protozoa); царство растений Plante. Домены включают царства, типы, классы, порядки, семейства, роды, виды.





Систематика – это наука, к/я решает вопросы клсф мк-в. Располагает всех мк по опред группам в соответствии с их сходными ФТ и ГТ пр!! Эти группы, объединяющие отдельных мк, наз ТАКСОНАМИ. Выделяют след таксономические единицы: вид (species) > род (genus) > семейство (familia) > порядок > класс > тип > царство (Procaryotae, Eucaryotae, Vira)
Вид – это …
Т.к. внутри вида т/же есть различия м/у мк, то выделяют ещё:
ШТАММ – объединяет мк одного и того же вида, но выделенные из опред источника и в опред время.
КЛОН – это культура мк, полученная из единичной клетки, к/я размножилась и дала начало целой популяции.
Когда образуется большое скопление клеток, то среди них могут быть клетки с отклонениями – биовары (биологические варианты), хемовары, фаговары, серовары (серологические варианты, имеющие разные антигенные свойства), резистенсвары.
Вид не является конечной единицей систематики. Внутри вида выделяют варианты микроорганизмов, отличающиеся отдельными признаками. Так, различают:
1) серовары (по антигенной структуре);
2) хемовары (по чувствительности к химическим веществам);
3) фаговары (по чувствительности к фагам);
4) ферментовары;
5) бактериоциновары;
6) бактериоциногеновары.
Бактериоцины – вещества, продуцируемые бактериями и губительно действующие на другие бактерии. По типу продуцируемого бактериоцина различают бактериоциновары, а по чувствительности – бактерициногеновары.
Для видовой идентификации бактерий необходимо знать следующие их свойства:
1) морфологические (форму и структуру бактериальной клетки);
2) тинкториальные (способность окрашиваться различными красителями);
3) культуральные (характер роста на питательной среде);
4) биохимические (способность утилизировать различные субстраты);
5) антигенные.
Номенклатура – список названий разл групп мк. Кажд мк им св назв в осн к/го положена бинарная система, т.е. название мк состоит из названия рода и вида (Staphilococcus aureus). Как правило, родовое название пишется с заглавной буквы, м.б. сокращённым, видовое – полностью.
Отличия прокариот от эукариот:
Ø у эукариот (грибы и простейшие) есть ядро с оболочкой и ядрышком, ЭПС, МХ, рибосомы, КГ и проч. Размножаются митозом
Ø у прокариот — ядра нет, НК плавает в ЦП, скручена в спираль. Есть рибосомы
· - ЦП окружена мембраной, за ней — клеточная стенка
· - размножаются бинарным делением
· - в клеточной стенке содержатся пептидогликаны, определяющие окраску по Граму (много Грам+, мало Грам-)
Прокариоты - это, как правило, одноклеточные организмы, бактерии, отличающиеся слабой морфологической дифференцировкой (доядерные); для них характерно:
· - отсутствие окруженного мембраной ядра (носителем наследственности является нуклеоид - замкнутая в кольцо нить ДНК, единственная "бактериальная хромосома");
· - отсутствие органелл (митохондрий, хлоропластов, комплекса Гольджи и др.);
· - размножение бинарным амитотическим делением (надвое);
· - особое строение и состав клеточной стенки, малые размеры рибосом, своеобразные ферменты белкового синтеза.
Микроскопические эукариоты - это относительно более высоко организованные одноклеточные и многоклеточные организмы, имеющие сходство с клетками животных (простейшие) и растений (грибы). Для эукариот характерны:
· - наличие истинного ядра, в котором находится набор линейных хромосом, распределяющихся в ходе митоза в дочерние клетки;
· - различные органеллы (митохондрии, комплекс Гольджи, эндоплазматический ретикулюм и др.);
· - рибосомы большего размера, чем у прокариот;
· - способность к эндоцитозу (захвату частиц и растворенных веществ).
Заключение
Учет результатов. Заключение по исследованию. Учитывают результаты идентификации и по совокупности полученных данных, опираясь на классификацию и характеристику типовых штаммов, описанных в руководстве (определитель Берджи, 1994-1996 гг.), определяют вид выделенных культур.
14. Вирусы. Классификация. Морфология и строение вирионов. Химический состав, функции структурных элементов. Прионы. Вироиды. Работы Д.И. Ивановского.









Морфология и ультраструктура вирусов бактерий - бактериофагов. Стадии взаимодействия вирулентного и умеренного фага с бактериальной клеткой. Лизогения. Фаговая конверсия. Практическое применение фагов.
Материальные основы наследственности микроорганизмов. Организация генетического материала у прокариотов, эукариотов, вирусов. Генотип и фенотип. Виды изменчивости. Модификации. Механизм. Фенотипические проявления.
Мигрирующие генетические элементы бактерий: транспозоны, is-последовательности, плазмиды. Характеристика. Свойства плазмид. Роль плазмид в детерминировании патогенных признаков и лекарственной устойчивости.



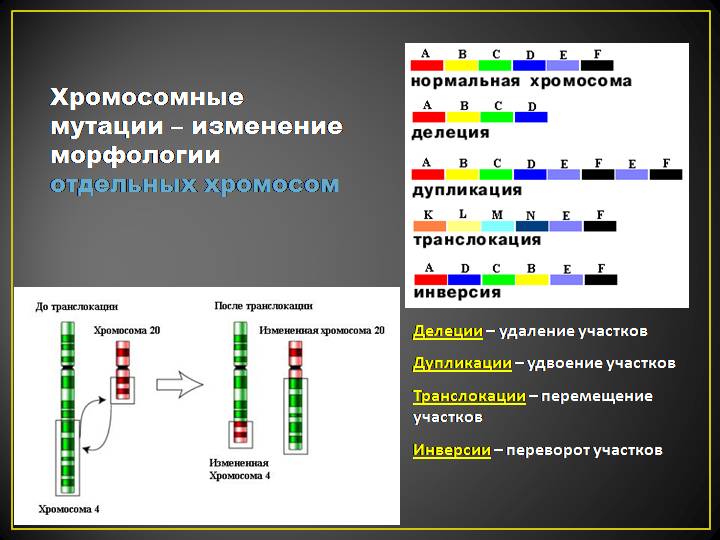
Мутации, их классификации. Мутагены физические, химические, биологические. Молекулярный механизм мутаций (делеция, дупликация, инверсия). Репарации и их значение. Роль мутаций в эволюционном процессе.
Репарация (от лат. reparatio — восстановление) — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических агентов. Осуществляется специальными ферментными системами клетки. Ряд наследственных болезней (напр., пигментная ксеродерма) связан с нарушениями систем репарации. (Процесс, позволяющий живым организмам восстанавливать повреждения, возникающие в ДНК, называют репарацией – кратко). Все репарационные механизмы основаны на том, что ДНК - двухцепочечная молекула, т.е. в клетке есть 2 копии генетической информации. Если нуклеотидная последовательность одной из двух цепей оказывается повреждённой (изменённой), информацию можно восстановить, так как вторая (комплементарная) цепь сохранена. Процесс репарации происходит в несколько этапов. На первом этапе выявляется нарушение комплементарности цепей ДНК. В ходе второго этапа некомплементарный нуклеотид или только основание устраняется, на третьем и четвёртом этапах идёт восстановление целостности цепи по принципу комплементарности. Однако в зависимости от типа повреждения количество этапов и ферментов, участвующих в его устранении, может быть разным. Очень редко происходят повреждения, затрагивающие обе цепи ДНК, т.е. нарушения структуры нуклеотидов комплементарной пары. Такие повреждения в половых клетках не репарируются, так как для осуществления сложной репарации с участием гомологичной рекомбинации требуется наличие диплоидного набора хромосом.
Виды: 1)Прямая репарация 2)Эксцизионная репарация 3)Пострепликативная репарация
· Прямая репарация — наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты, способные быстро (как правило, в одну стадию) устранять соответствующее повреждение, восстанавливая исходную структуру нуклеотидов. Так действует, например, O6-метилгуанин-ДНК-метилтрансфераза, которая снимает метильную группу с азотистого основания на один из собственных остатков цистеина. Эксцизионная репарация (англ. excision — вырезание) включает удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы.
· Эксцизионная репарация (excision repair): процесс с участием ферментативной системы, которая удаляет короткую однонитевую последовательность двунитевой ДНК, содержащей ошибочно спаренные или поврежденные основания, и замещает их путем синтеза последовательности, комплементарной оставшейся нити. Эксцизионная репарация является наиболее распространенным способом репарации модифицированных оснований ДНК. Этот тип репарации базируется на распознавании модифицированного основания различными гликозилазами, расщепляющими N-гликозидную связь этого основания с сахарофосфатным остовом молекулы ДНК. При этом существуют гликозилазы, специфически распознающие присутствие в ДНК определенных модифицированных оснований (оксиметилурацила, гипоксантина, 5-метилурацила, 3-метиладенина, 7-метилгуанина и т.д.). Для многих гликозилаз к настоящему времени описан полиморфизм, связанный с заменой одного из нуклеотидов в кодирующей последовательности гена. Для ряда изоформ этих ферментов была установлена ассоциация с повышенным риском возникновения онкологических заболеваний [ Chen, 2003 ].
· Пострепликативная репарация Tип репарации, имеющей место в тех случаях, когда процесс эксцизионной репарации недостаточен для полного исправления повреждения: после репликации с образованием ДНК, содержащей поврежденные участки, образуются одноцепочечные бреши, заполняемые в процессе гомологичной рекомбинации при помощи белка RecA.[1] Пострепликативная репарация была открыта в клетках E.Coli, не способных выщеплять тиминовые димеры. Это единственный тип репарации, не имеющий этапа узнавания повреждения.

Систематика микроорганизмов (прокариоты, грибы, простейшие, вирусы). Принципы классификации. Таксономические категории. Понятие о виде как основной таксономической единице. Номенклатура. Основные отличия прокариотов от эукариотов.
Микроорганизмы представлены доклеточными формами (вирусы — царство Vira) и клеточными формами (бактерии, архебактерии, грибы и простейшие). Различают 3 домена (или «империи»): «Bacteria», «Archaea» и «Eukarya»:
Ø □ домен «Bacteria» — прокариоты, представленные настоящими бактериями (эубактериями);
Ø □ домен «Archaea» — прокариоты, представленные архебактериями;
Ø □ домен «Eukarya» — эукариоты, клетки которых имеют ядро с ядерной оболочкой и ядрышком, а
цитоплазма состоит из высокоорганизованных органелл — митохондрий, аппарата Гольджи и др. Домен «Eukarya» включает: царство Fungi (грибы); царство животных Animalia (включает прстейшие – подцарство Protozoa); царство растений Plante. Домены включают царства, типы, классы, порядки, семейства, роды, виды.
Систематика – это наука, к/я решает вопросы клсф мк-в. Располагает всех мк по опред группам в соответствии с их сходными ФТ и ГТ пр!! Эти группы, объединяющие отдельных мк, наз ТАКСОНАМИ. Выделяют след таксономические единицы: вид (species) > род (genus) > семейство (familia) > порядок > класс > тип > царство (Procaryotae, Eucaryotae, Vira)
Вид – это …
Т.к. внутри вида т/же есть различия м/у мк, то выделяют ещё:
ШТАММ – объединяет мк одного и того же вида, но выделенные из опред источника и в опред время.
КЛОН – это культура мк, полученная из единичной клетки, к/я размножилась и дала начало целой популяции.
Когда образуется большое скопление клеток, то среди них могут быть клетки с отклонениями – биовары (биологические варианты), хемовары, фаговары, серовары (серологические варианты, имеющие разные антигенные свойства), резистенсвары.
Вид не является конечной единицей систематики. Внутри вида выделяют варианты микроорганизмов, отличающиеся отдельными признаками. Так, различают:
1) серовары (по антигенной структуре);
2) хемовары (по чувствительности к химическим веществам);
3) фаговары (по чувствительности к фагам);
4) ферментовары;
5) бактериоциновары;
6) бактериоциногеновары.
Бактериоцины – вещества, продуцируемые бактериями и губительно действующие на другие бактерии. По типу продуцируемого бактериоцина различают бактериоциновары, а по чувствительности – бактерициногеновары.
Для видовой идентификации бактерий необходимо знать следующие их свойства:
1) морфологические (форму и структуру бактериальной клетки);
2) тинкториальные (способность окрашиваться различными красителями);
3) культуральные (характер роста на питательной среде);
4) биохимические (способность утилизировать различные субстраты);
5) антигенные.
Номенклатура – список названий разл групп мк. Кажд мк им св назв в осн к/го положена бинарная система, т.е. название мк состоит из названия рода и вида (Staphilococcus aureus). Как правило, родовое название пишется с заглавной буквы, м.б. сокращённым, видовое – полностью.
Отличия прокариот от эукариот:
Ø у эукариот (грибы и простейшие) есть ядро с оболочкой и ядрышком, ЭПС, МХ, рибосомы, КГ и проч. Размножаются митозом
Ø у прокариот — ядра нет, НК плавает в ЦП, скручена в спираль. Есть рибосомы
· - ЦП окружена мембраной, за ней — клеточная стенка
· - размножаются бинарным делением
· - в клеточной стенке содержатся пептидогликаны, определяющие окраску по Граму (много Грам+, мало Грам-)
Прокариоты - это, как правило, одноклеточные организмы, бактерии, отличающиеся слабой морфологической дифференцировкой (доядерные); для них характерно:
· - отсутствие окруженного мембраной ядра (носителем наследственности является нуклеоид - замкнутая в кольцо нить ДНК, единственная "бактериальная хромосома");
· - отсутствие органелл (митохондрий, хлоропластов, комплекса Гольджи и др.);
· - размножение бинарным амитотическим делением (надвое);
· - особое строение и состав клеточной стенки, малые размеры рибосом, своеобразные ферменты белкового синтеза.
Микроскопические эукариоты - это относительно более высоко организованные одноклеточные и многоклеточные организмы, имеющие сходство с клетками животных (простейшие) и растений (грибы). Для эукариот характерны:
· - наличие истинного ядра, в котором находится набор линейных хромосом, распределяющихся в ходе митоза в дочерние клетки;
· - различные органеллы (митохондрии, комплекс Гольджи, эндоплазматический ретикулюм и др.);
· - рибосомы большего размера, чем у прокариот;
· - способность к эндоцитозу (захвату частиц и растворенных веществ).
Строение и химический состав клеточной стенки грамположительных и грамотрицательных бактерий. Бактерии с дефектом синтеза клеточной стенки: протопласты, сферопласты, L – формы.







Микроорганизмы относятся к царству прокариот: отсутствие ядерной мембраны, слабо развитая ЭПС, нет митохондрий, хлоропластов, нет КГ, не обладают эндоцитозом. Ядерное вещество представлено 1 хромосомой. Она двухнитчатая, кольцевидной формы молекулы ДНК. Ядерный материал сосредоточен в центральной части и имеет видфиломентозных образований. Цитоплазма имеет вид зернистого образования, куда входят различные вакуоли, рибосомы, гликоген, крахмал, зерна волютина (питательный запас).
ЦПМ состоит из 3 слоев, выполняет барьерную функцию, поддерживает осмотическое давление в клетке, способствует переходу питательных веществ внутрь клетки и выходу продуктов обмена веществ во внешнюю среду. Клеточная стенка у разных видов микроорганизмов различна по строению. В состав ее входит редкий гетерополимер- пептидогликан- у разных микроорганизмов он имеет различную толщину, на чем основан один из основных методов окраски- по Граму. В соответствии с окрашиванием микроорганизмы делятся на 2 группы: грам положительные и грам отрицательные. У Гр+ микроорганизмов в составе клеточной стенки- большое количество пептидогликана, он многослойный, на его долю приходится 80% от химического состава клеточной стенки. У Гр- пептидогликана всего 10% и он однослойный. Тейхоевые кислоты в стенке Гр+ имеются, у Гр- их нет. За счет этих 2 компонентов микроорганизмы окрашиваытся разно: Гр+- в синий цвет(удерживают генциан виолет). Гр- окрашиваются фуксином в красный цвет. Отношение к окраске - это тинкториальные свойства.
Клеточная стенка - это необязательный структурный элемент. Ее не имеют микоплазмы, L-формы микроорганизмов. L-формы образуются из обычных микроорганизмов, которые по ряду причин теряют клеточную стенку. Без клеточной стенки микроб более уязвим к факторам окружающей среды.
ЦПМ имеет специфические выпячивания внутрь цитоплазмы – мезосомы. Они делятся на центральные и латеральные. Мезосомы способны соединяться с ДНК и участвовать в делении бактериальной клетки- растаскивают нити ДНК по полюсам.
Многие микроорганизмы имеют капсулу, она состоит из скопления слизистого материала и выполняет защитную функцию. У патогенных микроорганизмов капсула образуется только внутри макроорганизма, она является фактором патогенности, препятствует фагоцитозу, доступу к микробу антител, бактериофагов. Капсула не окрашивается обычными красителями, только по Гимсу- Бурри. Препарат смешивают с черной тушью, которая образует темный фон, затем окрашивают фуксином.

4. Капсулы. Жгутики. Пили. Химический состав и функциональное значение этих образований у бактерий. Методы выявления. Патогенные представители.



5. Споры и спорообразование. Химический состав и функциональное значение спор. Методы выявления. Патогенные представители.

6. Метаболизм бактерий. Ферменты бактерий и их роль в обмене веществ. Конститутивные и индуцибельные, экзо- и эндо- ферменты. Практическое использование биохимической активности бактерий.

Ферменты являются высокоспецифичными биологическими катализаторами, без которых невозможны жизнь и размножение. Большое количество реакций, происходящих при жизни бактериальной клетки, указывает на существование у бактерий значительного количества ферментов. Ферменты — вещества белковой природы с большим молекулярным весом. Некоторые из них относятся к протеинам, другие являются сложными белками. Они построены из двух частей белка и небелковой части, называемой простетической группой. В состав ее могут входить витамины, нуклеотиды, атомы железа и пр. Связь между белковой частью фермента и его простетической группой может быть прочной и непрочной. При наличии непрочной связи в растворах наступает диссоциация фермента и при этом может освобождаться свободная простетическая группа. Легко диссоциирующие простетические группы ферментов называются коферментами. Обычно ферменты подразделяются на следующие основные группы:
1. Оксидоредуктазы - катализируют реакции окисления-восстановления.
2. Трансферазы - катализируют реакции переноса различных групп от донора к акцептору.
3. Гидролазы - катализируют разрыв связей в субстратах с присоединением воды.
4. Лиазы - катализируют реакции разрыва связей в субстрате без присоединения воды или окисления.
5. Изомеразы - катализируют превращения в пределах одной молекулы (внутримолекулярные перестройки).
6. Лигазы (синтетазы) - катализируют присоединение двух молекул с использованием энергии фосфатных связей.
Наиболее высокой ферментативной активностью обладают сапрофиты; в меньшей степени это свойство выражено у патогенных бактерий. Изучение ферментов патогенных бактерий имеет исключительно важное значение, так как на основании определения ферментативной активности микробов можно дифференцировать различные виды и определять природу того или иного возбудителя заболеваний. Наряду с этим ферментативная активность микробов определяет патогенез и клиническую картину инфекционного заболевания. Ферменты дифференцируют на экзо и эндоферменты.
Экзоферменты выделяются клеткой во внешнюю среду, осуществляют процессы расщепления высокомолекулярных органических соединений на более простые, доступные для ассимиляции.
Ферменты бактерий подразделяются на: конституитивные, индуцибельные.
· относятся те ферменты, которые синтезируются бактериальной клеткой вне зависимости от того, на какой среде бактерия выращивается. К ним относятся основные ферменты клеточного обмена — липазы, карбогидразы, протеиназы, оксидазы.
· Индуцибельные ферменты продуцируются данной бактерией лишь в ответ на действие специфического индуктора, присутствующего в среде. Например, р-галактозидаза кишечной палочкой на среде с глюкозой практически не образуется, но ее синтез резко увеличивается при выращивании палочек на среде с лактозой или другим галактозидом.
По своей локализации бактериальные ферменты подразделяются на две группы.
· Ферменты, которые синтезируются бактериальной клеткой в окружающую среду и действуют в околоклеточном пространстве, называются экзоферментами. К ним относятся, протеолитические ферменты грамположительных бактерий, пеницилиназы и т.п. Такая локализация ферментов обусловлена их функциональной задачей – они, расщепляют макромолекулярный питательный субстрат на низкомолекулярные части, способные транспортироваться внутрь клетки, или инактивируют вещества, обладающие антибактериальной активностью, «на подходе» к клетке.
· Ферменты, которые локализуются внутри бактериальной клетки, называются эндоферментами. Эндоферменты содержатся в периплазматическом пространстве грамотрицательных бактерий, а также в цитоплазматической мембране и цитоплазме.Б. По зависимости своего синтеза от наличия субстрата, на который действует фермент, бактериальные ферменты подразделяются также на две группы.
Известны также ферменты, которые получили название аллостерических. Кроме активного центра у них имеется регуляторный или аллостерический центр, который в молекуле фермента пространственно разделен с активным центром. Аллостерическим (от греч. allos - иной, чужой) он называется потому, что молекулы, связывающиеся с этим центром, по строению (стерически) не похожи на субстрат, но оказывают влияние на связывание и превращение субстрата в активном центре, изменяя его конфигурацию. Молекула фермента может иметь несколько аллостерических центров. Вещества, связывающиеся с аллостерическим центром, называют аллостерическими эффекторами. Они влияют через аллостерический центр на функцию активного центра: или облегчают ее, или затрудняют. Соответственно аллостерические эффекторы называются положительными (активаторы) или отрицательными (ингибиторы). Аллостерические ферменты играют важную роль в тонкой регуляции метаболизма бактерий.
Поскольку практически все реакции в клетке катализируются ферментами, регуляция метаболизма сводится к регуляции интенсивности ферментативных реакций.






