Взаимосвязь физики и биологии давняя и плодотворная. Можно назвать выдающихся физиков, внесших свой вклад в развитие биологии, и естествоиспытателей, открывшие фундаментальные физические законы. Это всемирно известные: физик Гельмгольц, врач Майер, ботаник К. А. Тимирязев и др. П. Н. Лебедев, приветствуя К. А. Тимирязева в связи с его семидесятилетием сказал: «Мы, физики, считаем Вас физиком!»
Однако преподавателей физики, так же, как и преподавателей биологии, должны интересовать не отдельные примеры проявления физических явлений и законов в живой природе, а система знаний, раскрывающих перед учащимися взаимосвязь и взаимообусловленность биологических и физических явлений. Важно показать учащимся, что установление этой взаимосвязи помогает глубже понять сущность сложных биологических явлений и найти пути не только их изучения, но и управления этими явлениями.
Можно указать следующие пути установления связи преподавания физики с биологией:
· Изучение физических явлений и закономерностей, определяющих физические условия внешней среды (тепловые, оптические и другие свойства почвы и приземного слоя воздуха), и влияние их на жизнь растений, животных и самого человека; сюда же относится изучение электромагнитных явлений и различных видов излучений, которые имеют место при земном слое атмосферы и оказывают влияние на органический мир;
· Ознакомление учащихся с физическими явлениями и закономерностями в биологических процессах роста и развития растений, а также в жизни животных и человека; к таким явлениям можно отнести испарение, фотосинтез, процессы теплообмена, передачи давления и другие;
· Изучение физических приборов и физических методов исследования, применяемых в биологии.
При рассмотрении примеров проявления физических явлений и законов в биологических процессах важно не только показывать единство живой и не живой природы, но и особенности физических явлений в биологических объектах.
Методы изучения физических явлений живой природы в процессе преподавания физики могут быть самыми разнообразными. Соответствующие сведения могут сообщаться учащимися в качестве иллюстративного материала. Так, при изучении молекулярно-кинетической теории строения вещества полезно рассказать о диффузии жидкостей и газов в природе.
При изучении электричества можно сообщить учащимся, что электрические заряды и электрическое поле играют большую роль в жизнедеятельности клеток. В невозбужденном стоянии клетки всегда заряжены снаружи положительно, а в нутрии отрицательно. Напряжение между внешней оболочкой и внутренними частями клеток составляет 0,05- 0,1 В.
Разность потенциалов возникает благодаря тому, что оболочка клетки неодинаково поглощает и пропускает различные ионы. На образование разности потенциалов расходуется энергия, освобождающаяся при обмене веществ.
В живых организмах постоянно существуют биотоки. Правда, биотоки в растениях и животных слабые. Например, у мелких корней растений сила тока равна всего 0,01 мкА.
С физическими свойствами биологических объектов, особенностями их строения или протекающими в них процессами можно познакомить учащихся с помощью демонстраций.
Ряд интересных сведений учащиеся могут получить при решении задач.
В организме человека происходит непрерывно окисление питательных веществ. «Горючим» является прежде сего углеводы и жиры и в меньшей мере – белки. При окислении в организме 1 грамма белков и углеводов даёт нам 17 Дж, а 1 грамм жиров – 38 Дж энергии. Сообщив эти данные, можно предложить учащимся следующую задачу: «Рассчитать калорийность суточного рациона для людей, выполняющих тяжелый физический труд (землекопы, лесорубы, шахтеры и т.д.), если они должны потреблять в среднем 163 граммов белков, 153 грамма жиров и 631 грамм углеводов».
Далее приведены части лекций курса физики, с раскрытием тем через биологические объекты и примеры:
1) Тема «Кинематика»
Человек и любые другие биологические объекты, являясь частью материального мира, естественно, подчиняются законам механики и физики. Вместе с тем живые объекты отличаются от технических систем и обладают рядом особенностей, которые следует иметь в виду, изучая их механическое поведение. Важнейшей особенностью является способность живых систем адаптироваться к длительным или многократным воздействиям внешних факторов путем функциональной и морфологической перестройки отдельных структур.
С точки зрения биомеханики аппарат движения человека представляет собой управляемую систему подвижно соединенных костей, обладающих определенными размерами, массами, моментами инерции и снабженных мышечными двигателями. Скелет состоит из 206 костей (85 парных и 36 непарных), которые соединены суставами и связками. Это пассивная часть опорно-двигательного аппарата.
Поперечнополосатые скелетные мышцы (их более 600) — это его активная часть, приводящая в движение костные звенья. Управляет этим костно-мышечным аппаратом движения центральная нервная система.
Само движение осуществляется системой костных рычагов, которые приводятся в движение силой тяги, возникающей при сокращении мышц.
Рычагом называется твердое тело, имеющее ось вращения (точку опоры), к которому приложены силы, создающие моменты сил относительно этой оси. Напомним, что момент силы (относительно некоторой оси или точки опоры) равен произведению величины силы и плеча ее действия, а плечо силы — это кратчайшее расстояние от оси вращения до линии действия силы, лежащей в плоскости вращения.
В качестве примера рычага рассмотрим предплечье (рис. 1.1, а). Здесь сила тяги мышцы F приложена к короткому плечу а рычага (точка опоры О — локтевой сустав), а к его длинному плечу b приложена нагрузка R — сила тяжести удерживаемого груза, приложенная обычно к кисти, а также сила тяжести самого предплечья, приложенная в его центре масс (на рисунке не показана и далее не учитывается). Моменты этих сил равны соответственно Fa и Rb.
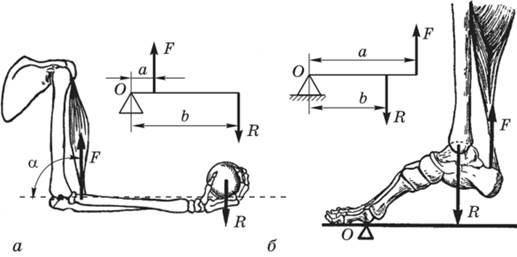
Рис. 1.1. Основные виды рычага: а — рычаг скорости; б — рычаг силы
Для равновесия рычага необходимо равенство моментов противодействующих сил, приложенных к нему:
 (1.1)
(1.1)
При движении рычага угловая скорость всех его точек  одинакова, поэтому для точек приложения груза и мышцы к рычагу можем записать
одинакова, поэтому для точек приложения груза и мышцы к рычагу можем записать
 (1.2)
(1.2)
где 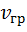 — скорость движения груза;
— скорость движения груза;  — скорость укорочения мышцы.
— скорость укорочения мышцы.
Из этих соотношений следует, что  .
.
При  рычаг проигрывает в силе (
рычаг проигрывает в силе ( ), но выигрывает в скорости перемещения груза и называется поэтому рычагом скорости (например, предплечье, рис. 1.1, а). При
), но выигрывает в скорости перемещения груза и называется поэтому рычагом скорости (например, предплечье, рис. 1.1, а). При  рычаг дает выигрыш в силе (
рычаг дает выигрыш в силе (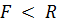 ) и называется рычагом силы, но он проигрывает в скорости перемещения груза. Примером такого рычага служит свод стопы при подъеме на полупальцы (рис. 1.1, б). В этом случае опорой О рычага служат головки плюсневых костей; преодолеваемая сила тяжести R тела приложена к таранной кости, а мышечная сила F, осуществляющая подъем тела, передается через ахиллово сухожилие и приложена к выступу пяточной кости и поскольку здесь
) и называется рычагом силы, но он проигрывает в скорости перемещения груза. Примером такого рычага служит свод стопы при подъеме на полупальцы (рис. 1.1, б). В этом случае опорой О рычага служат головки плюсневых костей; преодолеваемая сила тяжести R тела приложена к таранной кости, а мышечная сила F, осуществляющая подъем тела, передается через ахиллово сухожилие и приложена к выступу пяточной кости и поскольку здесь  .
.
Часто мышечная сила F направлена к рычагу под углом a, отличным от 90° (рис. 1.2). В этом случае условие равновесия рычага принимает вид:  , откуда
, откуда  , т.е. мышечное усилие F, необходимое для преодоления данной силы сопротивления R, должно быть тем больше, чем под меньшим углом к оси рычага оно направлено.
, т.е. мышечное усилие F, необходимое для преодоления данной силы сопротивления R, должно быть тем больше, чем под меньшим углом к оси рычага оно направлено.

Рис. 1.2. Вариант рычага скорости при  < 90°
< 90°
Одиночное мышечное волокно при сокращении развивает силу порядка 2 мН. В мышцах человека содержится примерно  волокон. Если бы все они сократились одновременно в одном направлении, то создали бы огромное усилие — до 60 000 Н.
волокон. Если бы все они сократились одновременно в одном направлении, то создали бы огромное усилие — до 60 000 Н.
Сила мышцы зависит от количества содержащихся в ней волокон. Поскольку их трудно подсчитать, обычно измеряют поперечное сечение мышцы, которое пропорционально числу волокон, и по нему судят о ее силовых возможностях. При этом нужно определять не геометрическое, а физиологическое поперечное сечение, под которым понимают сумму поперечных сечений всех волокон, образующих данную мышцу.
Отношение максимального мышечного усилия, которое может развить мышца, к ее физиологическому поперечному сечению называется абсолютной мышечной силой и выражается в единицах механического напряжения (Н • м2 = Па). Для большинства скелетных мышц человека абсолютная мышечная сила имеет порядок 106 Па (100 Н/см2), а для гладких — 105 Па (10 Н/см2). Сила одних и тех же мышц человека зависит от ряда физиологических условий: возраста, пола, тренированности и т.д.
Перемещение тела человека в пространстве совершается благодаря мышечным сокращениям, при которых мышцы укорачиваются и напрягаются. В изотоническом режиме, т.е. при постоянной нагрузке на мышцу, зависимость между скоростью и мышечного сокращения и приложенной силой F описывается уравнением Хилла:
 (1.3)
(1.3)
Где  — максимальное усилие, развиваемое данной мышцей; а — константа, имеющая размерность силы; b — константа, имеющая размерность скорости.
— максимальное усилие, развиваемое данной мышцей; а — константа, имеющая размерность силы; b — константа, имеющая размерность скорости.
Из уравнения Хилла (1.3) следует, что при отсутствии нагрузки (F = 0) скорость сокращения мышцы максимальная, с увеличением нагрузки она уменьшается и при максимальной нагрузке ( ) ее укорочение вовсе не происходит (у = 0), что демонстрирует и рис. 1.3.
) ее укорочение вовсе не происходит (у = 0), что демонстрирует и рис. 1.3.

Рис. 1.3. Зависимость скорости  укорочения мышцы от приложенной к ней нагрузки
укорочения мышцы от приложенной к ней нагрузки 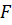
Коэффициент полезного действия (КПД= А/Е) при сокращении мышцы определяется как отношение совершенной работы А к затраченной энергии Е и может достигать 40 - 60 % у разных типов мышц.
Мышечные усилия и внешние силы, действующие на ткани человека, вызывают деформацию этих тканей и создают в них механические напряжения, которые до определенных значений безопасны для организма, но их превышение может приводить к травмам: растяжению мышц, связок, вывихам, переломам и др.
2) Тема «Работа и энергия»
В соответствии с законом Пуазейля течение жидкости по сосудам определяется разностью давлений в начале и конце сосудов. В системе кровообращения этот перепад давлений обусловлен в основном работой сердца. Таким образом, по отношению к сосудистой системе сердце можно считать некоторым насосом. В отличие от обычного поршневого насоса, где площадь поршня постоянна, при работе сердца его рабочая поверхность — внутренняя поверхность желудочков — изменяется в различные фазы их сокращения. Сила F, действующая на кровь в желудочках, зависит от давления Р в их полости и площади внутренней поверхности S:

В начале систолы давление меньше, а объем и площадь поверхности полости желудочков — больше. В конце систолы давление возрастает, а внутренняя поверхность уменьшается. Количественное рассмотрение этих процессов и непосредственные физиологические эксперименты показывают, что при уменьшении объема внутренних полостей сердце развивает меньшую силу. Согласно закону Старлинга, сила, развиваемая при сокращении сердца, пропорциональна начальной длине волокон миокарда.
Не занимаясь рассмотрением сложных явлений, происходящих при сокращении сердца, рассчитаем его работу за одно сокращение исходя из общих физических соображений. Эта работа А складывается из работы левого Ал и правого Ап желудочков:

Работа левого желудочка, выбрасывающего кровь в большой круг кровообращения, естественно, больше, чем работа правого желудочка. Считается, что, Ап = 0,2Ал. Таким образом А = 1,2АЛ.
Работа левого желудочка при выбросе систолического (ударного) объема крови в аорту затрачивается на преодоление сил давления крови в сосудистой системе и на сообщение крови кинетической энергии. Первая составляющая этой работы называется статическим компонентом, а вторая — кинетическим компонентом.
Статический компонент рассчитаем по формуле

где Р ср— давление крови в аорте; V c — систолический объем.
Считая для человека в покое Р ср = 100 мм рт. ст. = 13,3 кПа и V c = 60 мл = 6*10-5 м3, найдем А ст = 0,8 Дж.
Кинетический компонент может быть рассчитан из формулы для кинетической энергии

Где m — масса крови в ее систолическом объеме; р — плотность крови; v — линейная скорость крови при выбросе в аорту.
Приняв р = 1,05 103 кг/м3, и = 0,5 м/с, получим, что Ак= 0,008 Дж.
Таким образом, работа сердца за одно сокращение определяется по формуле
 (1.4)
(1.4)
В покое она составляет примерно 1 Дж. Считая, что в среднем сердце сокращается один раз в секунду, определим работу сердца за сутки. Она составляет 86 400 Дж. Для сравнения, такую работу надо совершить, чтобы поднять груз массой в 1 т на высоту около 9 м.
Учитывая, что продолжительность систолы составляет около 0,3 с, и разделив работу сердца за одно сокращение на это время, получим значение средней мощности сердца в покое — 3,3 Вт.
При физической нагрузке работа сердца резко увеличивается, возрастает систолический и минутный объем крови, увеличивается и скорость течения крови в аорте. При этом изменяется и соотношение между статическим и кинетическим компонентами. Как видно из приведенных численных значений, в покое доля кинетического компонента составляет около 1 % общей работы сердца. С повышением нагрузки растут оба слагаемых формулы (1.4), но кинетический компонент растет быстрее, так как он пропорционален квадрату увеличивающейся линейной скорости кровотока и доля кинетического компонента в общей работе сердца может достигать 30 %.
3) Тема «Термодинамика»
В живой системе работа не может совершаться за счёт притока тепла извне, как в тепловой машине, она совершается за счёт изменения внутренней энергии системы при различного рода биохимических процессах или за счёт энтропийного фактора.
Говоря образно: живая система – не паровоз, сколько таракана не подогревай, он за счёт этого тепла быстрее двигаться не будет.
В настоящее время не вызывает сомнения применение закона сохранения энергии к биологическим объектам. Однако так было не всегда. Первые экспериментальные попытки измерения энергетического баланса организма были предприняты в конце 18 в. А. Лавуазье и П. Лапласом. Для доказательства применимости первого начала термодинамики к биологическим объектам был применен ледяной калориметр. По скорости таяния льда находили теплоту, выделенную морской свинкой в изолированной камере или ледяном калориметре. После этого определяли количество теплоты, выделяющейся при прямом сжигании до СО2 и Н2О продуктов питания морской свинки в калориметрической бомбе. А. Лавуазье и П. Лаплас получили величины близких порядков.
Таким образом, метод калориметрии позволил установить, что количество энергии, поглощенной организмом за сутки вместе с питательными веществами, равно выделенной за это же время теплоте.
Энергетический баланс организма находится в полном соответствии с законом сохранения энергии, и никаких специфических источников жизни не существует.
Жизнедеятельность организмов, как и работа тепловой машины, сопровождается выделением в окружающую среду тепловой энергии за счет дыхания, пищеварения, работы мышц.
Удельной теплопродукцией называют количество теплоты, выделяемое единицей массы животного в единицу времени:

где Q t - количество теплоты, выделяемое в единицу времени; m - масса; g - удельная теплопродукция.
Теплопродукция пропорциональна массе организма. Удельная теплопродукция уменьшается с увеличением массы. Такую зависимость можно объяснить тем, что с увеличением размера и массы животного уменьшается отношение его поверхности к объему. Для компенсации большой потери теплоты мелкими животными требуется более высокая интенсивность метаболизма и, соответственно, потребление большего количества пищи. Этим можно объяснить необычайную прожорливость грызунов и мелких птиц.
Живые организмы - открытые термодинамические системы, существующие в условиях постоянства давления и температуры. Поэтому для оценки их жизнедеятельности важен еще один термодинамический параметр (функция) - энтальпия.
Энтальпия (от греч. - нагреваю) — это та энергия, которая доступна для преобразования в теплоту при определенных температуре и давлении.
Рассчитывается по формуле:

где Н - энтальпия, U - внутренняя энергия, P - давление, V - объём.
Энтальпия - термодинамическая функция, которая, подобно температуре, давлению, объему и внутренней энергии, характеризует систему. Абсолютное значение энтальпии для рассматриваемой системы определить невозможно и в термодинамических расчетах фигурирует лишь изменение энтальпии ΔН, происходящее при переходе системы из одного состояния в другое.
Физический смысл введения понятия энтальпии: изменение энтальпии — это тепло, подведенное к системе в изобарическом процессе (при постоянном давлении). Практическое применение этой функции основано на том, что множество химических процессов в реальных или лабораторных условиях реализуются именно при постоянном (атмосферном) давлении, когда резервуар открыт. Так, энтальпия образования сложного вещества - количество энергии, которое выделяется или поглощается при образовании сложного вещества из простых веществ.
Все химические реакции сопровождаются выделением (экзотермические) или поглощением (эндотермические) тепла. Мерой теплоты реакции служит изменение энтальпии ΔН, которая соответствует теплообмену при постоянном давлении. В случае экзотермических реакций система теряет тепло и ΔН – величина отрицательная. В случае эндотермических реакций система поглощает тепло и ΔН – величина положительная.
Введенная функция лежит в основе Закона Гесса – основного следствия первого закона термодинамики. Он формулируется следующим образом: тепловой эффект химической реакции, имеющей ряд промежуточных стадий, не зависит от пути, а определяется лишь разностью энтальпий конечных и исходных продуктов реакции.
Известно, что биохимические превращения в организме проходят множество промежуточных стадий, которые не всегда выявлены и доказаны. На основании закона Гесса, не зная промежуточных звеньев биохимического процесса, имея данные лишь о начальных и конечных продуктах реакции, можно вычислить суммарный энергетический эффект.
Закон Гесса используется для вычисления калорийности пищевых продуктов и составления пищевых рационов человека и животных.
4) Тема «Второе начало термодинамики»
Первое начало термодинамики не дает ответа на вопрос о том, почему процесс, связанный с трансформацией энергии, протекает именно в данном направлении; насколько полно осуществляется превращение энергии; каковы реальные пределы протекания того или иного биохимического процесса.
Второе начало термодинамики позволяет судить о направлении протекания процесса и, таким образом, дополняет первое начало термодинамики.
Формулировку второго начала термодинамики впервые дали независимо друг от друга Р. Клаузиус и У. Томпсон. Р. Клаузиус: "Теплота не может переходить сама собой от более холодного тела к более теплому".
Второе начало не имеет столь простой и общепринятой формулировки, как первое, и формулируется по-разному, в зависимости от той группы явлений, к которой оно прилагается.
Важной физической величиной, используя которую можно сформулировать второе начало термодинамики в более общем виде, является еще один термодинамический параметр систем – энтропия.
Энтропия является таким же свойством термодинамической системы, как температура, давление и др. В любом теле (лист бумаги) содержится определенная энтропия; так же, как и внутренняя энергия, энтропия системы растет с ее массой и равна сумме энтропии подсистем, она увеличивается при повышении температуры за счет усиления теплового движения молекул. Аналогичное увеличение энтропии наблюдается при переходе вещества из твердого состояния в жидкое. Для вычисления изменения энтропии ΔS биохимической реакции из суммы изменения энтропии продуктов реакции вычитают сумму изменения энтропии исходных веществ.
Изменение энтропии ΔS системы равно отношению количества теплоты (dQ), сообщенного системе, к температуре (Т):

Если ввести понятие энтропии, то второе начало термодинамики можно сформулировать так: энтропия изолированной системы возрастает в необратимом процессе и остается неизменной в обратимых термодинамических процессах. Уравнение второго закона термодинамики:

Сущность второго начала термодинамики для изолированных систем состоит в том, что все необратимые процессы протекают в таком направлении, в котором энтропия системы увеличивается.
Энтропия — это часть общей энергии клетки, которая не может быть использована в данной системе. Согласно второму началу термодинамики энтропия изолированной системы стремится к максимуму, при котором достигается равновесие, и реакция прекращается. Если атомы в молекуле упорядочены, то энтропия системы низка.
Второй закон термодинамики показывает, что не все виды энергии эквивалентны. По качеству их можно разделить на 3 класса (А, В, С) соответственно величине энтропии. Такие виды энергии, как гравитационная, световая и электрическая, наиболее эффективно, т. е. с наименьшей энтропией, могут быть превращены в другие ее виды, и они относятся к классу А. В класс В (среднего качества) попадает химическая энергия. Тепловая энергия относится к классу С (низкое качество) — это энергия с максимальной энтропией.
Живые организмы - системы открытые, и изменение энтропии для них складывается из продукции энтропии внутри организма за счет необратимых биохимических процессов и обмена энтропией с окружающей средой.
Формулировка второго закона термодинамики для живых организмов: скорость изменения энтропии в организме равна алгебраической сумме производства энтропии внутри организма и скорости поступления энтропии из среды в организм.
Биологические процессы могут сопровождаться самопроизвольным уменьшением энтропии. Так, рост и развитие организмов сопровождается усложнением их организации и с точки зрения классической термодинамики выглядит как самопроизвольное уменьшение энтропии и увеличение (накопление) свободной энергии. Дело в том, что свободная энергия не может возрастать лишь в изолированной системе. Живые организмы, как системы открытые, в процессе автотрофного и гетеротрофного питания получают свободную энергию из окружающей среды. Поэтому второй закон термодинамики применительно к живым организмам надо рассматривать, учитывая их взаимодействие с окружающей средой. С этой точки зрения уменьшение энтропии и увеличение свободной энергии фотосинтезирующих организмов происходит благодаря уменьшению свободной энергии и увеличению энтропии в системе Солнце-Земля (ядерные реакции на Солнце) и общий баланс энтропии автотрофов соответствует второму началу термодинамики.
Точно так же уменьшение энергии в частях клетки, где идет биохимический синтез, происходит за счет избыточного увеличения энтропии в реакциях диссимиляции, и общий баланс энтропии увеличивается. Поэтому живые организмы подчиняются второму началу термодинамики, когда речь идет о круговороте и превращениях свободной энергии в системе организм - окружающая среда.
Состояние системы, при котором ее параметры не изменяются в течение длительного времени, но происходит обмен веществом и энергией с окружающей средой, называют стационарным. В стационарном состоянии открытой системы концентрация промежуточных продуктов не изменяется со временем, что достигается определенным соотношением различных физико-химических процессов, ответственных за распад и образование промежуточных соединений.
Живой организм в каждый момент времени не отвечает приведенному определению стационарного состояния. Однако, если рассмотреть средние значения его параметров за сравнительно большой промежуток времени, можно отметить их постоянство и утверждать, что стационарное состояние характерно для организма. Так, с большим постоянством поддерживается температура определенных органов и тканей у теплокровных животных, сохраняется неизменный солевой состав и водородный показатель (рН) различных биологических жидкостей, не изменяются в ходе нормальной жизнедеятельности величины биопотенциалов покоя, осмотического давления.
В открытых системах суммарное изменение энтропии в стационарном состоянии равно нулю.
Стационарное состояние биологической системы поддерживается, когда увеличение энтропии системы за счет происходящих в ней необратимых процессов компенсируется оттоком энтропии при взаимодействии с окружающей средой.
Особенности стационарного состояния и термодинамического равновесия:
- cходство термодинамического равновесия и стационарного состояния системы заключается в том, что стационарное состояние, так же, как и термодинамическое равновесие, сохраняет все основные параметры неизменными.
- энтропия системы, находящейся в стационарном состоянии, имеет некоторую постоянную величину, не равную максимальной. Поэтому наиболее характерными свойствами стационарного состояния являются стремление системы к минимуму ежесекундного прироста энтропии и определенная внутренняя стабильность, и упорядоченность.
В организме поддерживается гомеостаз в течение времени, протекающего от рождения до смерти. Уровни стационарных состояний в течение онтогенеза постоянно изменяются. У детей уровень Са2+ в костях с их ростом увеличивается, изменяется количество зубов, волос и т. д. Это примеры перехода на новые стационарные уровни, примеры необратимых изменений.
Живой организм может изменить уровень стационарного состояния в результате воздействия окружающей среды и при патологических процессах.
Одной из важнейших характеристик биологических систем является устойчивость стационарных состояний. Устойчивое стационарное состояние характеризуется тем, что при отклонении системы от стационарного уровня в ней возникают силы, стремящиеся вернуть ее в первоначальное положение.
Внешние воздействия вызывают в неустойчивой стационарной системе нарастающие изменения, в результате которых система переходит или в устойчивое стационарное состояние (при дополнительной затрате энергии), или в состояние термодинамического равновесия.

Рис. 1.4 Диаграмма устойчивого (а) и неустойчивого (b) стационарных состояний в открытой системе
Если построить график зависимости Т∙dS/dt от h (где Т - температура, dS/dt - скорость производства энтропии, h - какой-либо показатель стационарного уровня системы), то для устойчивого стационарного состояния график будет представлен в виде параболы, ветви которой направлены вверх. При неустойчивом стационарном состоянии ветви параболы направлены вниз. Точка А является наиболее устойчивой. Если же поместить шарик во внутрь параболы, то его положение наиболее устойчиво. Таким образом, устойчивое стационарное состояние характеризуется тем, что система не может самопроизвольно выйти из него за счет внутренних изменений (шарик из точки А не может скатиться в точку В самопроизвольно, для этого нужно совершить работу). Точка Ai - наиболее неустойчивое состояние системы, так как под влиянием любого внешнего толчка шарик, помещенный в эту точку, быстро удаляется от нее.
Любое отклонение от уровня стационарности вызовет в системе увеличение скорости продуцирования энтропии. Для точки А скорость производства энтропии будет меньше, чем для точки В.
Система, которая функционирует в одном из двух устойчивых стационарных состояний, характеризуется триггерными свойствами, обеспечивающими направленные и скачкообразные переключения из одного стационарного состояния в другое. Например, при раздражении нервного волокна состояние возбуждения, возникающее по принципу "все или ничего", наступает скачкообразно после достижения порога возбуждения. Триггерные свойства биологических систем определяют и процесс клеточной дифференциации, когда при делении возникают дочерние клетки, качественно отличные от клеток-предшественников.
Теорема Пригожина
И. Пригожин (1946) на основе изучения открытых систем сформулировал основное свойство стационарного состояния: в стационарном состоянии при фиксированных внешних параметрах скорость продукции энтропии в открытой системе, обусловленная протеканием необратимых процессов, постоянна во времени и минимальна по величине.
Таким образом, согласно теореме Пригожина, стационарное состояние характеризуется минимальным рассеянием энергии.
По теореме Пригожина для поддержания стационарного состояния тратится некоторое количество энергии, причем в каждом случае самое минимальное. Организм стремится работать на самом выгодном энергетическом режиме. При этом энтропия возрастает с минимальной скоростью.
Однако теорема Пригожина не позволяет характеризовать стационарное состояние с точки зрения устойчивости к внешним воздействиям и самопроизвольным флуктуациям в самой открытой системе. Теорема Пригожина приложима к живым организмам лишь в качестве первого приближения, ибо она предполагает постоянство кинетических параметров, которое в организме в полной мере не выдерживается, особенно в начальный период его развития.
Стационарное состояние живого организма характеризуется высокой динамичностью: в организме протекают такие процессы, которые нарушают стационарное состояние тех или иных систем (повышение температуры тела при воспалительных процессах). Если эти изменения не превышают определенных пределов, то стационарное состояние может восстановиться, т. е. небольшие отклонения от стационарного состояния не ведут к необратимым изменениям. Биологические системы обладают способностью к саморегуляции (установление наиболее экономичных, наиболее надежных уровней в работе живых организмов). Это положение впервые было сформулировано в работе Ле-Шателье.
Всякая система, находящаяся в состоянии химического равновесия и отклонившаяся от этого состояния под воздействием внешнего возмущения, стремится самопроизвольно вернуться в равновесное состояние за счет изменения параметров в направлении, противоположном тому, которое вызвало возмущение.
Принцип автостабилизации биологических систем имеет прямое отношение к теореме Пригожина, так как стремление энтропии в стационарном состоянии к минимальной величине приводит к тому, что при отклонениях от стационарного уровня в системе наступают такие изменения, которые стремятся вернуть ее к минимуму производства энтропии. Принцип Ле-Шателье является следствием закона сохранения энергии, лежит в основе регулирования по типу обратных связей и поддержания гомеостаза в организме.
Принцип автостабилизадии в применении к биологическим системам состоит в том, что увеличение потока положительной энтропии при отклонении системы от стационарного уровня (заболевание) приводит к появлению таких внутренних изменений, которые ограничивают этот поток, т. е. в организме происходят процессы, способствующие ликвидации причин, вызывающих заболевание, и система настраивается на более термодинамически выгодный ритм функционирования.
5) Тема «Жидкости»
Поверхностное натяжение определяет форму биологических клеток и их частей. Изменение сил поверхностного натяжения влияет на фагоцитоз (захват клетками соседних частиц), пиноцитоз (захват клеточной поверхностью жидкости с содержащимися в ней веществами), на процессы альвеолярного дыхания.
Поверхностное натяжение различных веществ колеблется от 0,01 до 2,0 Н/м, у клеток оно не превышает 0,10 Н/м. Вещества, растворенные в жидкости, способны как понижать, так и повышать поверхностное натяжение. Вещества, понижающие поверхностное натяжение раствора, называются поверхностно-активными веществами (ПАВ). К ним относится, в частности, сурфактант, снижающий поверхностное натяжение альвеолярных стенок, обеспечивая тем самым возможность дыхания.
6) Тема «Механические колебания и волны»
Человеческое ухо и глаз являются наиболее удивительными и эффективными устройствами, созданными когда-либо для восприятия информации, переносимой волнами. В случае уха падающая звуковая волна связывается различными его участками с чувствительными нервами, которые ассоциируются со слухом. Рис. 1.5 показывает наружную часть уха человека. Звук поступает в систему через ушной канал, который является закрытой с одной стороны акустической трубой с длиной примерно 2,7 см.
Столб воздуха внутри ушного канала будет резонировать на звук с длиной волны, равной четырем его длинам. Основная резонансная частота v = v/(3L) = 3 кГц. Этот резонансный эффект отчасти объясняет тот факт, что человеческое ухо наиболее чувствительно в диапазоне частот приблизительно 3 кГц.
В конце ушного канала находится барабанная перепонка. Изменения давления падающей звуковой волны передаются барабанной перепонкой системе небольших костей (косточек) в среднем ухе (рис. 1.6). Эти кости называются в соответствии с их формой молоточек, наковальня и стремя. Косточки служат для передачи отклика внешнего уха внутреннему, или улитке. Среднее ухо заполнено воздухом. Любая разность давлений между наружным и средним ухом приводит к деформации барабанной перепонки.
Таким образом, звуковая информация передается костям среднего уха. Однако любое заметное изменение давления окружающего воздуха (например, связанное с изменением высоты), действуя на перепонку, может вызвать боль или даже ее разрыв. Для избавления от таких перепадов давления служит небольшая трубка (евстахиева труба), которая соединяет полость наружного уха с верхней частью глотки. Евстахиева труба, с одной стороны, достаточно велика, что позволяет проходить воздуху и приспосабливаться к большим изменениям давления, и, с другой стороны, достаточно мала, чтобы быстрые изменения давления, вызванные звуковыми волнами, могли быть переданы через барабанную перепонку. (Некоторы




