Аннотация
Во время 1- гоЗа тысячелетие до нашей эры кочевые племена, связанные с скифской культурой железного века, распространились по Евразийской степи, охватывая территорию шириной более 3500 км. Чтобы понять демографические процессы, стоящие за распространением скифской культуры, мы проанализировали геномные данные восьми особей и митохондриальный набор данных 96 особей, происходящих из восточной и западной частей Евразийской степи. Геномный вывод показывает, что скифы на востоке и западе степной зоны лучше всего можно описать как смесь родства, связанного с Ямной, и восточноазиатского компонента. Демографическое моделирование предполагает независимое происхождение для восточных и западных групп с постоянным потоком генов между ними, правдоподобно объясняя поразительную однородность их материальной культуры.
Вступление
В течение первого тысячелетия до нашей эры кочевые народы распространялись по Евразийской степи от Алтайских гор до северного Причерноморья, вплоть до Карпатского бассейна 1. Классические скифы, жившие в регионе Северного Причерноморья с седьмого века до нашей эры, являются наиболее известными среди них благодаря ранним сообщениям в «Историях Геродота» (490 / 480–424 до н.э.) 2. Греческие и персидские историки 1- гоТысячелетие до нашей эры ведет хронику существования Massagetae и Sauromatians, а затем, Sarmatians и Sacae: культуры, обладающие артефактами, подобными найденным в классических скифских памятниках, таких как оружие, упряжь для лошадей и отличительная художественная традиция «Животного стиля». Соответственно, эти группы часто относятся к скифской культуре и называются «скифами». Для упрощения мы будем использовать «скифский» в следующем тексте для всех групп степных кочевников железного века, обычно связанных с скифской культурой.
Происхождение широко распространенной скифской культуры уже давно обсуждается в евразийской археологии. Северное Причерноморье вначале считалось родиной и центром скифов 3,пока Тереножкин не сформулировал гипотезу о центральноазиатском происхождении 4. С другой стороны, доказательства, подтверждающие восточно-евразийское происхождение, включают в себя курган Аржан 1 в Туве 5, который считается самым ранним скифским курганом 5. Датирование дополнительных мест захоронений, расположенных на востоке и западе Евразии, подтвердило, что восточные курганы старше своих западных коллег 6, 7. Кроме того, элементы характерного «звериного стиля» датируются X веком до н.э. 1, 4были обнаружены в районе реки Енисей и современного Китая, что способствовало раннему присутствию скифской культуры на Востоке. Артефакты скифской культуры распространились по большой территории вскоре после ее появления, но основная динамика численности населения, которая, возможно, стала причиной культурного распространения, плохо изучена.
Генетические исследования популяций Центральной Азии, основанные как на древней 8, так и на современной митохондриальной ДНК (мтДНК) 9, 10, 11, 12, подтверждают, что Центральная Азия исторически была перекрестком для перемещения населения с востока на запад и наоборот. Утверждалось, что поток генов происходил с востока на запад Евразии еще в эпоху палеолита 13, 14 и мезолита 15, а также с запада на восток Евразии во время бронзового века 16. Недавнее геномное исследование 17подчеркнул роль миграций людей на восток, связанных с ямной и андроновской культурой в эпоху бронзы, с последующим существенным смешением с восточноазиатскими народами. Однако большинство генетических исследований поздних кочевников железного века были ограничены небольшим размером выборки, ограничены одной культурной группой или основаны на анализе только одной мтДНК 18, 19, 20, 21, 22.
В этом исследовании были секвенированы и проанализированы данные мтДНК от 96 особей, связанных со скифской культурой в разных географических регионах и периодах времени; Кроме того, были получены и проанализированы геномные данные восьми из этих лиц (дополнительная таблица 1).). Из западной части Евразийской степи обнаружены образцы, обнаруженные на Северном Кавказе в начале скифского периода (восьмой-шестой век до н.э.), классические скифы из Доно-Поволжья (третий век до н.э.) и ранние сарматы из Покровки на юго-западе. Урала (с пятого по второй век до н.э.), были включены. Из восточной части Евразийской степи мы проанализировали образцы из Восточного Казахстана, датируемые фазой Зевакино-Чиликта (с девятого по седьмой век до н.э.); с сайта Аржан 2, отведенного культуре Алды Бел в Туве (с седьмого по шестой век до н.э.); и из тагарской культуры Минусинского бассейна (V век до н.э.). Большинство образцов, полученных в результате этого исследования или извлеченных из литературы, относятся к IV-III векам до н.э. и были обнаружены в археологических памятниках, расположенных на казахском языке, Русская и Монгольская части Алтайских гор. Все эти находки были приписаны пазырыкской культуре, которая названа в честь первых открытий Грязнова в 1927 и 1929 годах в Пазырыкской долине и известна своими богатыми морозоустойчивыми могилами, где хорошо сохранились человеческие тела, гобелен и одежда.23, 24, 25 (рис. 1).
Рисунок 1: Реконструкция скифа.

Найден в кургане Олон-Курин-Гол 10, Алтайские горы, Монголия (реконструкция Дмитрия Позднякова).
В то время как восточное и западное население, исследуемое здесь, разделено расстоянием 2000–3500 км, археологические данные указывают на то, что они были поразительно похожи в отношении их образа жизни и культуры. Цели данного исследования состоят в том, чтобы исследовать, насколько эти группы генетически связаны друг с другом и имеют ли они общее происхождение, и выяснить их демографическую историю и генетические связи с современным живым населением. Поэтому мы разделили данные о степных кочевниках железного века, полученные в результате этого исследования и из литературы (n = 147, дополнительная таблица 1), на семь выборочных групп (см. Рис. 2).), основываясь на географических и хронологических критериях, и проанализировал этих древних особей вместе с обширной выборкой современных особей из 86 популяций (n = 3410, дополнительная рис. 11 и дополнительная таблица 19) из всех частей Евразии. Наш анализ включал в себя множество статистических методов, а также ряд популяционных подходов генетического вывода, включая явное демографическое моделирование.
Рисунок 2: Распределение участков выборки, проанализированных для этого исследования (желтый), включая данные из литературы (зеленый).
Числа относятся к определенным группам (#): ЗАПАД: # 1 начальный скифский период с восьмого по шестой век до н.э. (n = 4); # 2 классическая скифская фаза с VI по II век до н.э. (n = 19); # 3 Ранние сарматы с пятого по второй век до н.э. (n = 11); ВОСТОК: фаза Зевакино-Чиликта № 4 до н.э. до VII в. До н.э. (n = 11); # 5 культура Алды Бел в седьмом-шестом веке до нашей эры (n = 15); # 6 пазырыкская культура IV-III вв. До н.э. (n = 71); # 7 Тагар / Тесская культура восьмого века до н.э. - первого века нашей эры (n = 16); стрелки с буквой G обозначают образцы, для которых были получены геномные данные, черные для данных захвата и серые для данных о дробовике. Источник карты: картомедиа-Карлсруэ.
Результаты
Таблица 1: Список всех древних особей, из которых ядерные данные были получены в этом исследовании.
Мы дополнительно получили однозначные и воспроизводимые последовательности мтДНК гипервариабельной области 1 (HVR1; bp 16,013–16,410) для 96 образцов (из 110 образцов, для которых был предпринят анализ) (дополнительная таблица 5). Для образцов 90/96 были введены дополнительные SNP области кодирования (дополнительная таблица 6).
Рисунок 3: Возможные сценарии происхождения скифских поселений.
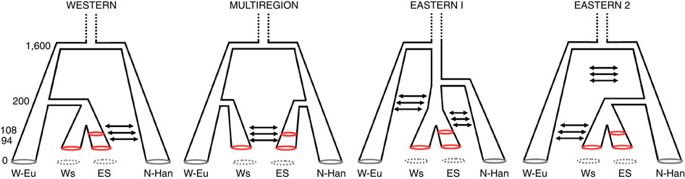
W-Eu = западные евразийцы; WS = западно-скифские группы; ES = восточно-скифские группы; N-Ha n = ханьский китайский. Числа слева относятся к поколениям до настоящего времени.
Поскольку перемещения населения через Центральную Азию во время бронзового века часто археологически связаны с распространением андроновской культуры 29, мы использовали ABC, чтобы соответствовать образцу кочевых групп среднего бронзового века из Западной Сибири, большинство из которых связаны с андроновской культурой, на предпочтительная демографическая модель происхождения скифов. Для этой цели - и на основании низких значений F ST между этими группами - мы объединили 40 образцов, относящихся к андроновской культуре в лесной степи Западной Сибири, и 30 образцов из той же культуры в Красноярском крае 31, причем все они были датированы первая половина 2- готысячелетие до нашей эры. Результаты обеспечили очень сильную поддержку связи между этими группами среднего бронзового века и восточными скифами (дополнительные таблицы 16 и 17). Тем не менее, эти модели не смогли полностью охватить модели генетического разнообразия, наблюдаемые в популяциях бронзового века, что позволяет предположить, что истинная демографическая история происхождения популяций железного века, возможно, была более сложной, чем рассматриваемая здесь (см. Дополнительные примечания 1 и 32). для деталей).
Рисунок 4: Анализ основных компонентов.

СПС древних особей (по цветам см. Легенду) проецируется на современных западноевразийцев (серый цвет). Скифы железного века изображены черным; CHG, охотник-собиратель Кавказа; LNBA, поздний неолит / бронзовый век; MN, средний неолит; EHG, восточноевропейский охотник-собиратель; LBK_EN, ранний неолитический линейный оркестр; HG, охотник-собиратель; EBA, ранний бронзовый век; IA, железный век; LBA, поздний бронзовый век; WHG, западный охотник-собиратель.
Поскольку СПС Западной Евразии на рис. 4 не позволяет исследовать древние образцы в отношении современных популяций Восточной Азии, мы также провели СПК всех 2345 современных особей в наборе данных «Происхождение человека» 35, на который мы также спроектировали древние особи (рис. 5). Из этого PCA очевидно, что происхождение образцов железного века приходится на континуум между современными западными евразийцами и восточными неафриканцами, что согласуется с анализом митохондриальной гаплогруппы. Восточные скифы демонстрируют почти равные пропорции линий мтДНК, распространенных в восточной и западной Евразии, тогда как в западных скифских группах частота линий, в настоящее время распространенных в восточной Евразии, как правило, ниже, даже достигая нуля в четырех образцах начальной скифской фазы с восьмого по шестое столетие до н.э. (группа № 1 на рис. 2) и достигает 18–26% в более поздние периоды (с шестого по второе столетие до н.э.; № 2 и № 3) (дополнительная таблица 7).
Рисунок 5: Анализ основных компонентов.
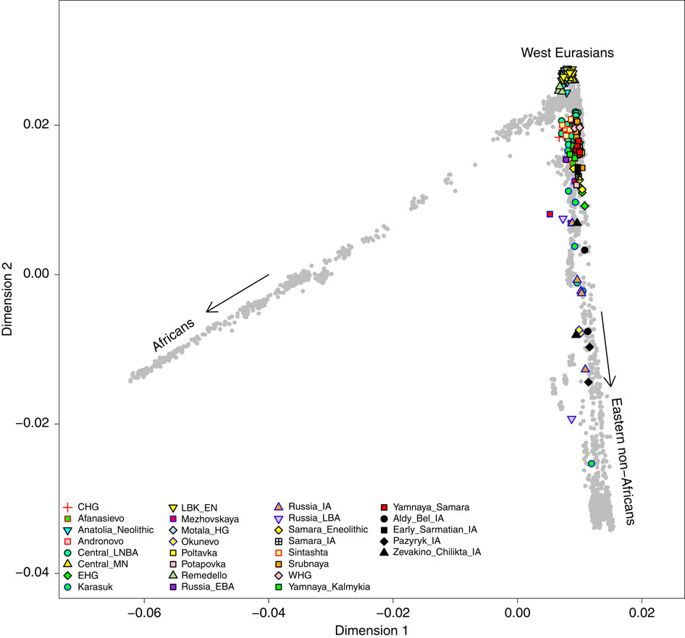
PCA древних людей (согласно цветам см. Легенду) проецируется на современных людей из набора данных о происхождении человека (серый). Скифы железного века изображены черным; CHG, охотник-собиратель Кавказа; LNBA, поздний неолит / бронзовый век; MN, средний неолит; EHG, восточноевропейский охотник-собиратель; LBK_EN, ранний неолитический линейный оркестр; HG, охотник-собиратель; EBA, ранний бронзовый век; IA, железный век; LBA, поздний бронзовый век; WHG, западный охотник-собиратель.
f -статистика
Мы использовали f 4-статистику вида f 4 (Test, LBK; EHG, Mbuti) и f 4 (Test, LBK; Han, Mbuti), которые равны нулю для тех тестовых образцов, которые образуют кладу с LBK и положительны для популяций. которые имеют происхождение, связанное с EHG или Ханом, соответственно. Мы нанесли результаты друг на друга, что привело к V-образному рисунку с Ямной на вершине (рис. 6). Скифы железного века расположены вдоль клина от Ямной до Ами (популяция восточноазиатских предков, в которой не было примеси), что соответствует происхождению от популяций, генетически сходных с этими двумя группами.
Рисунок 6: Визуализация результатов f- статистики.

Значения f 4 (Test, LBK; Han, Mbuti) нанесены на ось x, а значения f 4 (Test, LBK; EHG, Mbuti) на оси y, положительные отклонения от нуля показывают отклонения от поля между Test и LBK. Между Ямной из Самары и Ами проведена красная пунктирная линия. Популяции железного века, которые можно смоделировать как смеси ямной и восточно-евразийской (например, ами), расположены вокруг этой линии и, по-видимому, отличаются от основного северно-южноевропейского клина (синего цвета) слева от оси х.
Мы также вычислили статистику в форме f 3 (Test; Yamnaya_Samara, Han), чтобы проверить, имеет ли тестовая популяция промежуточные частоты аллелей между Yamnaya_Samara и Han, которые используются в качестве прокси для возможных исходных популяций. Промежуточные частоты аллелей могут иметь место только в том случае, если тестируемая популяция представляет собой смесь популяций, связанных с этими двумя источниками 37. Эти статистические данные значительно отрицательны для всех скифов, что свидетельствует о наличии примеси (дополнительная рис. 13).
ADMIXTURE анализ
Мы провели анализ ADMIXTURE 38, 39 из 2345 современных людей 35,генотипированных на массиве «Происхождение человека», 35, 37 и 175 древних особей на наборе из 296 340 SNP, пересекающихся с таковыми в массиве «Происхождение человека». Результаты для древних особей показаны на рис. 7 для K = 15, который имеет наибольшее логарифмическое значение правдоподобия (полный анализ можно найти на дополнительной рис. 14). Все степные популяции имеют компоненты предков, которые максимальны у европейских охотников-собирателей (синий) и кавказских охотников-собирателей из Грузии 36(зеленый). Одна подгруппа степных популяций (включая Срубную, Синташту и Андроново) также имеет раннюю фермерскую родословную (оранжевая), в то время как другая подгруппа (включая все образцы железного века) также имеет родословную от компонента (светло-голубой), который максимизируется в нганасане. (Самоеды из Северной Сибири), и распространены среди разнообразных современных людей из Сибири и Центральной Азии. Кроме того, в образцах железного века обнаружен наследственный компонент, который максимально увеличен в популяциях Восточной Азии (желтый), тип родословной, который встречается на уровне следов - если вообще вообще - среди более ранних обитателей степи, в соответствии с наблюдениями от PCA и f -statistics. об этом типе примеси.
Рисунок 7: Результаты ADMIXTURE для древних популяций.

Красные стрелки указывают на изученных скифских особей железного века. LBK_EN: ранний неолитический линейный бандкерамик; EHG: восточноевропейский охотник-собиратель; Motala_HG: охотник-собиратель из Моталы (Швеция); WHG: западный охотник-собиратель; CHG: охотник-собиратель Кавказа; IA: железный век; EBA: ранний бронзовый век; LBA: поздний бронзовый век.
Фенотипические маркеры
Производные аллели пигментных маркеров, которые отбираются у европейцев, присутствуют у восточных и западных скифов, в том числе у лиц, которые гомозиготны по производным аллелям в отобранных SNP в HERC2, SLC24A5 или SLC45A2 (ссылки 43, 44, 45). В двух локусах LCT, связанных с персистенцией лактазы, производный аллель наблюдается только у гетерозигот, только в образцах восточных скифов и с низкой частотой (2–3%). Аллельные предки в ADH1B rs3811801 и rs1229984 почти фиксированы в скифском наборе данных, как и у современных европейцев (производные аллели, которые придают некоторую устойчивость к алкоголизму, отбираются в Восточной Азии)46, 47). Заметимпроизводный аллель в rs3827760 в ГДДИ гена в одном пазырыкского индивидуума (# 6 нарис. 2). Этотпроизводный от EDAR аллель, связанный с морфологией зубов и волос, выбирается и с высокой частотой у современных восточноазиатских (87%) 48 и очень редко встречается у современных европейцев (∼ 1%) 48, хотя он наблюдался у доисторических охотники-собиратели из Швеции (7,9–7,5 тыс. лет) 34. Таким образом, результаты изучения фенотипических SNP, которые показывают различия в частоте между Европой и Восточной Азией, согласуются с потоком генов через территорию степи.
Обсуждение
Наши результаты показывают, что группы железного века, которые, как считается, связаны между собой общими культурными артефактами, связанными с классическими скифами Северного Причерноморья, также имеют генетическую связь. Это подтверждается нашим анализом ABC, показывающим непрерывность численности населения в 1- м тысячелетии до н.э. у восточных скифов и низкий F STценности между восточными и западными скифскими группами. Тем не менее, анализ ABC, в ходе которого оценивались различные модели происхождения скифских популяций, обеспечил наиболее сильную поддержку многорегионального происхождения, причем восточные и западные группы возникли независимо в своих регионах. Несмотря на раздельное происхождение и огромное географическое разделение, демографическое моделирование предполагает постоянный и значительный поток генов между восточными и западными группами, что обеспечивает правдоподобный демографический механизм, объясняющий низкие значения F ST и общую однородность материальной культуры скифов по всей Евразии. Степная зона.
Наш геномный анализ показывает, что западные и восточные степные жители в разной степени имеют восточно-евразийское происхождение. В наших анализах ADMIXTURE мы находим компонент родословной в Восточной Азии при K = 15 во всех образцах железного века, который не был обнаружен в предшествующих популяциях бронзового века ни в западной, ни в восточной частях Евразийской степи. Другой наследственный компонент, который максимизируется в северной сибирской популяции нганасан, становится видимым со 2- готысячелетие до н.э. в восточной степи (Окунево, Карасук, Межовская). Этот компонент появляется позже во всех популяциях железного века, но со значительно более высокими уровнями в восточной степной зоне, чем на западе. Эти данные согласуются с появлением восточно-евразийских линий митохондрий у западных скифов во время железного века и предполагают поток генов или миграцию по евразийскому степному поясу, несущему восточноазиатское / северо-сибирское происхождение от востока до запада вплоть до Доно-Поволжье на юге России. В целом, поток генов между восточной и западной Евразией, по-видимому, был более интенсивным в течение железного века, чем в современную эпоху, что согласуется с представлением о населении железного века Евразийской степи как о очень подвижной полукочевой верховой езде. групп.
На Востоке мы находим сбалансированную смесь митохондриальных линий, встречающихся сегодня преимущественно в западноевразийских странах, включая значительную долю доисторических линий охотников-собирателей и линий, которые встречаются с высокой частотой в современной Центральной и Восточной Азии уже у самых ранних особей железного века датируется девятым-седьмым веками до н.э. и еще более ранним образцом мтДНК из бронзового века в Монголии 49. Типичные линии мтДНК Западной Евразии также присутствуют в Таримском бассейне 16 и Казахстане 8 и даже преобладали в Красноярском крае в течение 2- го тысячелетия до н.э. 31, Эта модель указывает на процесс смешивания между западным и восточным евразийским населением, который начался в более ранние периоды, безусловно, до 1- готысячелетия до н.э. 13, 50, что согласуется с недавним исследованием, предполагающим, что носители ямной культуры генетически неотличимы от Афанасьево. культура народов Алтай-Саянского края. Это также означает, что носители ямной культуры мигрировали не только в Европу 26, но и на восток, неся западные евразийские гены - и, возможно, также индоевропейские языки - в этот регион 17, Все эти наблюдения свидетельствуют о том, что распространенный генетический паттерн не просто следует модели изоляции на расстоянии, но включает значительный поток генов на большие расстояния.
Все люди железного века, исследованные в этом исследовании, показывают геномные данные о происхождении охотников-собирателей Кавказа и стран Восточной Европы. Это согласуется с идеей, что смесь ЭХГ и кавказских элементов в носителях ямной культуры сформировалась в европейской степи и экспортировалась в Среднюю Азию и Сибирь 26. Все наши анализы подтверждают гипотезу о том, что генетический состав скифов может быть лучше всего описан как смесь родственных элементов Ямной и восточноазиатских / северносибирских элементов.
Что касается наследия кочевников железного века, мы находим, что современные человеческие популяции, имеющие тесную генетическую связь с скифскими группами, преимущественно расположены в непосредственной географической близости к отобранным местам захоронений, что указывает на определенную степень непрерывности популяции в исторические времена. Современные потомки западно-скифских групп встречаются среди различных групп на Кавказе и в Центральной Азии, в то время как сходства с восточными скифами обнаруживаются более широко, но почти исключительно среди тюркоязычных (бывших) кочевых групп, особенно из кипчакской ветви тюркской языки (дополнительное примечание 1). Генеалогическая связь между восточными скифами и носителями тюркского языка требует дальнейшего изучения, в частности, поскольку расширение тюркских языков считалось намного более новым, то есть, начиная с шестого века нашей эры, и происходило в процессе расширения элиты. Потенциально, есть много других демографических факторов, связанных с происхождением носителей тюркского языка, таких как волны миграции, связанные с сюнну, древнетюркским или ранним монгольским населением. Степень, в которой восточные скифы были вовлечены в раннее формирование тюркоязычных популяций, может быть выяснена будущими геномными исследованиями исторических периодов, следующих за скифскими временами.
Методы
Образец материала
Для этого исследования были отобраны образцы человеческих скелетов из разных частей России и Казахстана на основе связи археологического комплекса со скифскими погребальными обрядами и артефактами. Мы проанализировали 110 образцов скелета. Пять образцов не дали ДНК, восемь дали только плохие результаты, и для одной только кодирующей области можно было получить SNP. В итоге мы могли использовать 96 образцов для анализа мтДНК. Восемь человек были преобразованы в библиотеки Illumina; шесть из них были использованы для геномного захвата, два для секвенирования ружья и один для захвата и секвенирования ружья. Общая сохранность ДНК была на удивление хорошей, с небольшими вариациями в зависимости от региона или участка образца (дополнительные таблицы 1 и 2).
Базовые приготовления
Все этапы подготовки пробы перед ПЦР проводились в чистом помещении, физически отделенном от лабораторий после ПЦР. Подготовка образца, экстракция ДНК, амплификация отдельных фрагментов мтДНК и амплификация ядерных локусов были выполнены, как описано ранее 45. Для анализа мтДНК из литературы было отобрано 30 фрагментов кодирующих областей, охватывающих 32 специфичных для гаплогруппы SNP митохондриального генома 51,52, 53. Системы грунтовки (дополнительная таблица 3) были разработаны с использованием PrimerSelect, части программного пакета Lasergene (DNASTAR). Амплификация была выполнена в трех различных мультиплексных реакциях, чтобы избежать наложения фрагментов (дополнительная таблица 3). Мультиплексные ПЦР проводили в общем объеме 40 мкл, используя 20 мкл Qiagen Multiplex Kit Master Mix (Qiagen, Hilden, Германия), 11 мкл ddH 2 O, 0,2 мкл каждого праймера и 4 мкл экстракта ДНК. Условия мультиплексной ПЦР были следующими: начальная денатурация в течение 15 мин при 95 ° С с последующим 36 циклами денатурации 40 с при 94 ° С, отжиг 40–90 с при 56 ° С, удлинение 40–90 с при 72 ° С и для В некоторых реакциях добавляли конечное удлинение в течение 10 минут при 72 ° С. Стандартная ПЦР была использована для фрагментов, которые не смогли амплификации в мультиплексных реакциях и для амплификации HVR1 некоторых дополнительных образцов.
Биостатистический анализ
Для популяционного генетического анализа были использованы последовательности HVR1 из положения 16,040–16,400 из 96 образцов, проанализированных для этого исследования, и были использованы дополнительные 51 образец, взятый из литературы (дополнительная таблица 1). Программное обеспечение 59 Arlequin 3.5.1.3 использовалось для AMOVA и для расчета разнообразия генов (разнесения гаплотипов) 60, разнесения нуклеотидов 60, 61, значений F ST и расстояния Рейнольдса 62. Метод попарного расстояния использовался для расчета значений F ST с 1000 перестановками и значением гаммы = 0. Фу F S тест селективной нейтральности был выполнен для проверки расширения популяции. Уровень значимости для FЗначения S были установлены на уровне 0,02 (ссылка 63). Результаты сводной статистики см. В дополнительной таблице 7.
ABC анализ
Для изучения демографической истории скифов мы сформулировали несколько сценариев-кандидатов, которые послужили основой для моделирования выборок генетических данных для региона HVR-1 с использованием BayeSSC 64, 65. Расчеты сводной статистики по наблюдаемым данным были выполнены в DNaSP v5 (ссылка 66). Чтобы подтвердить, что сценарии-кандидаты смогли воспроизвести наблюдаемые генетические данные, мы сравнили предыдущие распределения смоделированной суммарной статистики с эмпирически наблюдаемыми значениями. Все анализы были выполнены с использованием пакета abc в R 2.15.1 (ссылки 67, 68). Мы ссылаемся на дополнительное примечание 1 для получения полной информации о демографических сценариях и анализах, используемых здесь.
Геномный анализ
Подготовка библиотеки ДНК для последующего секвенирования ружья проводилась в соответствии с протоколом, использованным в Kircher 2012 (ссылка 69), с небольшими изменениями для образцов ружья (дополнительное примечание 3). Для анализа геномных данных были вычислены f 3- и f 4 -статистики (qp3Pop и qpDstat из ADMIXTOOLS) 37, был применен анализ потоков предков (qpWave / qpAdm) 26 и был выполнен анализ ADMIXTURE 38, 39 (дополнительное примечание 2),
Доступность данных
Файлы BAM для геномных данных можно загрузить из Европейского архива нуклеотидов под регистрационным номером PRJEB18686. Митохондриальные последовательности были депонированы в GenBank под регистрационными номерами KY369766-KY369861.
Дополнительная информация
Как цитировать эту статью: Unterländer, M. et al. Родословная и демография и потомки кочевников железного века Евразийской степи. Туземный Commun. 8, 14615 дои: 10.1038 / ncomms14615 (2017).
Примечание издателя: Springer Nature остается нейтральным в отношении юрисдикционных претензий на опубликованных картах и институциональных принадлежностей.
Аннотация
Во время 1- гоЗа тысячелетие до нашей эры кочевые племена, связанные с скифской культурой железного века, распространились по Евразийской степи, охватывая территорию шириной более 3500 км. Чтобы понять демографические процессы, стоящие за распространением скифской культуры, мы проанализировали геномные данные восьми особей и митохондриальный набор данных 96 особей, происходящих из восточной и западной частей Евразийской степи. Геномный вывод показывает, что скифы на востоке и западе степной зоны лучше всего можно описать как смесь родства, связанного с Ямной, и восточноазиатского компонента. Демографическое моделирование предполагает независимое происхождение для восточных и западных групп с постоянным потоком генов между ними, правдоподобно объясняя поразительную однородность их материальной культуры.
Вступление
В течение первого тысячелетия до нашей эры кочевые народы распространялись по Евразийской степи от Алтайских гор до северного Причерноморья, вплоть до Карпатского бассейна 1. Классические скифы, жившие в регионе Северного Причерноморья с седьмого века до нашей эры, являются наиболее известными среди них благодаря ранним сообщениям в «Историях Геродота» (490 / 480–424 до н.э.) 2. Греческие и персидские историки 1- гоТысячелетие до нашей эры ведет хронику существования Massagetae и Sauromatians, а затем, Sarmatians и Sacae: культуры, обладающие артефактами, подобными найденным в классических скифских памятниках, таких как оружие, упряжь для лошадей и отличительная художественная традиция «Животного стиля». Соответственно, эти группы часто относятся к скифской культуре и называются «скифами». Для упрощения мы будем использовать «скифский» в следующем тексте для всех групп степных кочевников железного века, обычно связанных с скифской культурой.
Происхождение широко распространенной скифской культуры уже давно обсуждается в евразийской археологии. Северное Причерноморье вначале считалось родиной и центром скифов 3,пока Тереножкин не сформулировал гипотезу о центральноазиатском происхождении 4. С другой стороны, доказательства, подтверждающие восточно-евразийское происхождение, включают в себя курган Аржан 1 в Туве 5, который считается самым ранним скифским курганом 5. Датирование дополнительных мест захоронений, расположенных на востоке и западе Евразии, подтвердило, что восточные курганы старше своих западных коллег 6, 7. Кроме того, элементы характерного «звериного стиля» датируются X веком до н.э. 1, 4были обнаружены в районе реки Енисей и современного Китая, что способствовало раннему присутствию скифской культуры на Востоке. Артефакты скифской культуры распространились по большой территории вскоре после ее появления, но основная динамика численности населения, которая, возможно, стала причиной культурного распространения, плохо изучена.
Генетические исследования популяций Центральной Азии, основанные как на древней 8, так и на современной митохондриальной ДНК (мтДНК) 9, 10, 11, 12, подтверждают, что Центральная Азия исторически была перекрестком для перемещения населения с востока на запад и наоборот. Утверждалось, что поток генов происходил с востока на запад Евразии еще в эпоху палеолита 13, 14 и мезолита 15, а также с запада на восток Евразии во время бронзового века 16. Недавнее геномное исследование 17подчеркнул роль миграций людей на восток, связанных с ямной и андроновской культурой в эпоху бронзы, с последующим существенным смешением с восточноазиатскими народами. Однако большинство генетических исследований поздних кочевников железного века были ограничены небольшим размером выборки, ограничены одной культурной группой или основаны на анализе только одной мтДНК 18, 19, 20, 21, 22.
В этом исследовании были секвенированы и проанализированы данные мтДНК от 96 особей, связанных со скифской культурой в разных географических регионах и периодах времени; Кроме того, были получены и проанализированы геномные данные восьми из этих лиц (дополнительная таблица 1).). Из западной части Евразийской степи обнаружены образцы, обнаруженные на Северном Кавказе в начале скифского периода (восьмой-шестой век до н.э.), классические скифы из Доно-Поволжья (третий век до н.э.) и ранние сарматы из Покровки на юго-западе. Урала (с пятого по второй век до н.э.), были включены. Из восточной части Евразийской степи мы проанализировали образцы из Восточного Казахстана, датируемые фазой Зевакино-Чиликта (с девятого по седьмой век до н.э.); с сайта Аржан 2, отведенного культуре Алды Бел в Туве (с седьмого по шестой век до н.э.); и из тагарской культуры Минусинского бассейна (V век до н.э.). Большинство образцов, полученных в результате этого исследования или извлеченных из литературы, относятся к IV-III векам до н.э. и были обнаружены в археологических памятниках, расположенных на казахском языке, Русская и Монгольская части Алтайских гор. Все эти находки были приписаны пазырыкской культуре, которая названа в честь первых открытий Грязнова в 1927 и 1929 годах в Пазырыкской долине и известна своими богатыми морозоустойчивыми могилами, где хорошо сохранились человеческие тела, гобелен и одежда.23, 24, 25 (рис. 1).
Рисунок 1: Реконструкция скифа.
Найден в кургане Олон-Курин-Гол 10, Алтайские горы, Монголия (реконструкция Дмитрия Позднякова).
В то время как восточное и западное население, исследуемое здесь, разделено расстоянием 2000–3500 км, археологические данные указывают на то, что они были поразительно похожи в отношении их образа жизни и культуры. Цели данного исследования состоят в том, чтобы исследовать, насколько эти группы генетически связаны друг с другом и имеют ли они общее происхождение, и выяснить их демографическую историю и генетические связи с современным живым населением. Поэтому мы разделили данные о степных кочевниках железного века, полученные в результате этого исследования и из литературы (n = 147, дополнительная таблица 1), на семь выборочных групп (см. Рис. 2).), основываясь на географических и хронологических критериях, и проанализировал этих древних особей вместе с обширной выборкой современных особей из 86 популяций (n = 3410, дополнительная рис. 11 и дополнительная таблица 19) из всех частей Евразии. Наш анализ включал в себя множество статистических методов, а также ряд популяционных подходов генетического вывода, включая явное демографическое моделирование.
Рисунок 2: Распределение участков выборки, проанализированных для этого исследования (желтый), включая данные из литературы (зеленый).
Числа относятся к определенным группам (#): ЗАПАД: # 1 начальный скифский период с восьмого по шестой век до н.э. (n = 4); # 2 классическая скифская фаза с VI по II век до н.э. (n = 19); # 3 Ранние сарматы с пятого по второй век до н.э. (n = 11); ВОСТОК: фаза Зевакино-Чиликта № 4 до н.э. до VII в. До н.э. (n = 11); # 5 культура Алды Бел в седьмом-шестом веке до нашей эры (n = 15); # 6 пазырыкская культура IV-III вв. До н.э. (n = 71); # 7 Тагар / Тесская культура восьмого века до н.э. - первого века нашей эры (n = 16); стрелки с буквой G обозначают образцы, для которых были получены геномные данные, черные для данных захвата и серые для данных о дробовике. Источник карты: картомедиа-Карлсруэ.
Результаты
Образцы и данные последовательности
Мы сгенерировали данные захвата всего генома на целевом наборе из 1 233 553 SNP 26, 27 для шести особей: двух ранних сарматов из южного уральского региона (PR9, PR3, группа № 3 на рис. 2; пятый-второй век до н.э.), двух особи из Береля в Восточном Казахстане (Be9, Be11, # 6), относящиеся к пазырыкскому периоду (с четвертого по третий век до н.э.), и два человека, найденные в кургане Аржан 2 (A10, A17, # 5), приписанные к культуре Алды Бел в Туве (с седьмого по шестой век до н.э.). Для Be9 и двух дополнительных особей из восточного Казахстана (Is2 и Ze6, # 4), относящихся к фазе Зевакино-Чиликта (BCE от девятого до седьмого века), мы сгенерировали наборы данных полного генома с низким охватом (<0,3x) с помощью дробовика NGS (Таблица 1)., дополнительные таблицы 20 и 21).
Таблица 1: Список всех древних особей, из которых ядерные данные были получены в этом исследовании.
Мы дополнительно получили однозначные и воспроизводимые последовательности мтДНК гипервариабельной области 1 (HVR1; bp 16,013–16,410) для 96 образцов (из 110 образцов, для которых был предпринят анализ) (дополнительная таблица 5). Для образцов 90/96 были введены дополнительные SNP области кодирования (дополнительная таблица 6).








