Главной функцией мозжечка явл. координация и конроль сложных автоматизированных движений.
Выделяют 3 структуры мозжечка: древний (сост. из клочка, узелка и язычка червя), старый мозжечок включает центральную дольку, вершину, пирамиду, язычок червя и четырехугольную дольку полушарий. К новому мозжечку относятся полушария и часть червя (скат, листок и бугор).
Клетки Пуркинье (грушевидные нейроны), образующие средний слой коры, являются главной функциональной единицей с ярко выраженной интегративной направленностью. Она реализуется сильно ветвящимися дендритами, на которых в одной клетке может быть до 100 тыс. синапсов. Клетки Пуркинье являются единственными эфферентными нейронами коры мозжечка, непосредственно связывая ее с внутримозжечковыми и вестибулярными ядрами.
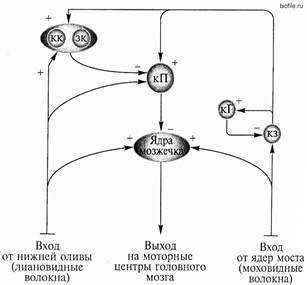 1) Почти все лиановидные волокна являются аксонами нейронов нижней оливы продолговатого мозга. Информация к ним поступает от мышечных, кожных рецепторов, а также от двигательной коры головного мозга. Каждое лиановидное волокно устанавливает контакты с дендритами, как правило, одной клетки Пуркинье (медиатор аспартат), но число синапсов может достигать трехсот, что объясняет исключительно сильное возбуждающее действие этого афферентного входа (большой ВПСП, на вершине которого формируется 3 — 5 ПД). Кроме того, лиановидные волокна оказывают на клетки Пуркинье и тормозное (более слабое) действие через корзинчатые и звездчатые клетки поверхностного слоя коры мозжечка.
1) Почти все лиановидные волокна являются аксонами нейронов нижней оливы продолговатого мозга. Информация к ним поступает от мышечных, кожных рецепторов, а также от двигательной коры головного мозга. Каждое лиановидное волокно устанавливает контакты с дендритами, как правило, одной клетки Пуркинье (медиатор аспартат), но число синапсов может достигать трехсот, что объясняет исключительно сильное возбуждающее действие этого афферентного входа (большой ВПСП, на вершине которого формируется 3 — 5 ПД). Кроме того, лиановидные волокна оказывают на клетки Пуркинье и тормозное (более слабое) действие через корзинчатые и звездчатые клетки поверхностного слоя коры мозжечка.
2) По моховидным волокнам в кору мозжечка поступает информация от коры больших полушарий (преимущественно ассоциативной), проприорецепторов, вестибулярных рецепторов и ретикулярной формации. Моховидные волокна образуют возбуждающие синапсы на дендритах клеток-зерен внутреннего слоя коры. Через аксоны клеток-зерен моховидные волокна оказывают непосредственное возбуждающее влияние (медиатор глутамат) на клетки Пуркинье и опосредованно тормозное действие через корзинчатые и звездчатые клетки. Активность клеток-зерен регулируется через тормозные клетки Гольджи внутреннего слоя по типу возвратного торможения (медиатор ГАМК).
3) Моноаминергический афферентный вход. Через него на мозжечок передается влияние голубого пятна (медиатор норадреналин) и ядер шва (медиатор серотонин). Стимуляция этих ядер тормозит фоновую активность клеток Пуркинье и модулирует их ответы на активацию моховидных и лиановидных афферентов.
На клетки Пуркинье поступает преимущественное возбуждающее афферентное влияние. Но, поскольку клетки Пуркинье являются тормозными нейронами (медиатор ГАМК), их возбуждение оказывает тормозное влияние на три парных мозжечковых ядра (ядра шатра, промежуточные и зубчатые), иннервирующих моторные центры головного мозга. Таким образом, мозжечок осуществляет регуляцию не по механизму запуска нейронной активности моторных центров, а по механизму более или менее сильного сдерживания этой активности.
Двигательные ф-и мозжечка. Регуляция мыш. тонуса, обеспечивающего поддержание естественной позы (равновесия) осущ. древним и частично старым мозжечком. Получая и обрабатывая импульсацию от вестиб. рецепторов, проприорецепторов аппарата движения, кожных, зрительных и слуховых рецепторов, мозжечок способен оценить состояние мышц, положение тела в пространстве и через ядра шатра, а также прямого выхода на вестибулярные ядра изменить позу тела и сохранить равновесие. Нарушение равновесия – статическая атаксия является наиболее характерным симптомом поражения древнего мозжечка.
Координация движений осущ. старым и новым мозжечком. В кору этой части мозжечка поступает импульсация от рецепторов аппарата движения, а также импульсация от моторной коры. Анализируя информацию о программе и выполнении движения, мозжечок способен через свои промежуточные ядра, имеющие выходы на красное ядро и моторную кору, осуществить координацию, а при необходимости и коррекцию движений. Нарушение координации движения (динамическая атаксия) является наиболее характерным симптомом нарушения функции промежуточной зоны мозжечка.
- асинергия – нарушение согласованности деятельности мышц-анатгонистов;
- адиадохокинез – затруднение в быстрой смене противоположных движений.
Программирование произвольных движений осуществляется корой нового мозжечка, которая получает информацию о замысле движения из ассоциативных зон коры большого мозга через ядра моста. В коре нового мозжечка она перерабатывается в программу движения, которая через зубчатое ядро мозжечка и вентральное латеральное ядро таламуса поступает в премоторную кору. Там она получает дальнейшую обработку и через пирамидную и экстрапирамидную системы реализуется как сложное целенаправленное движение (праксис, устная и письменная речь, нарушения которых являются одним из симптомов поражении нового мозжечка).
 17. Таламус – коллектор афферентных путей. Функциональная характеристика ядер таламуса, их роль в интегративной деятельности мозга.
17. Таламус – коллектор афферентных путей. Функциональная характеристика ядер таламуса, их роль в интегративной деятельности мозга.
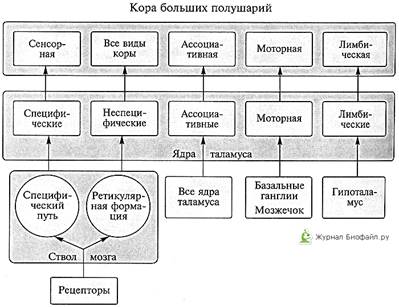
Таламус - парный ядерный комплекс, включающий до 60 ядер, в котором выделяют три главные группы ядер: релейные (специф., переключательные), ассоциативные и неспецифические. Все группы ядер таламуса тремя общими функциями: переключающей, интегративной и модулирующей.
1) Релейные ядра таламуса разделяют на сенсорные и несенсорные. Сенсорные ядра переключают потоки афферентной (чувствительной) импульсации в сенсорные зоны коры. В них также происходит перекодирование и обработка информации. Выделяют три основные ядерные структуры.
Вентральные задние ядра — главное реле для переключения соматосенсорной афферентной системы, импульсы которой поступают по волокнам медиальной, спинномозговой и тройничной петлям. В них переключаются тактильная, проприоцептивная, вкусовая, висцеральная, частично температурная и болевая чувствительность. В этих ядрах имеется топографическая проекция периферии; при этом функционально более тонко организованные части тела (например, язык, лицо) имеют большую зону представительства. Импульсация из вентральных задних ядер проецируется в соматосенсорную кору постцентральной извилины (поля 1—3), где форм.соответствующие ощущения. Электростимуляция вентральных задних ядер вызывает парестезии (ложные ощущения) в разных частях тела, иногда нарушение «схемы тела» (искаженное восприятие частей тела).
Латеральное коленчатое тело является реле для переключения зрительной импульсации в затылочную кору (поле 17), где она используется для формирования зрительных ощущений.
Медиальное коленчатое тело является реле для переключения слуховой и, возможно, вестибулярной импульсации в височную кору.
Переключательную функцию в сенсорных ядрах обеспечивают таламокортикальные (релейные) нейроны, длинный аксон которых непосредственно иннервируют нейроны сенсорной коры. Особенностью этой передачи является ее высокая точность с минимальным искажением входного сигнала.
Несенсорные релейные ядра таламуса (передние и вентральные переключают в кору несенсорную импульсаиию. поступающую в таламус из разных отделов головного мозга. В передние ядра импульсация поступает в основном из мамиллярных тел гипоталамуса. Нейроны передних ядер проецируются в лимбическую кору. От нее аксонные связи идут к гиппокампу и опять к гипоталамусу, в результате чего образуется нейронный круг, движение возбуждения по которому обеспечивает формирование эмоций. В связи с этим передние ядра таламуса рассматриваются как часть лимбической системы.
Вентральные ядра участвуют в регуляции движения, выполняя моторную функцию. В них переключается импульсация от базальных ядер, зуб. ядра мозжечка, красного ядра среднего мозга. После этого она проецируется в моторную и премоторную кору (поля 4 и 6). Через эти ядра таламуса происходит передача в моторную кору сложных двиг. программ, образованных в мозжечке и базальных ядрах.
2) Ассоциативные ядра таламуса принимают импульсацию от других ядер таламуса. Эфферентные выходы от этих ядер направляются в ассоциативные поля коры. В свою очередь кора мозга посылает волокна к ассоциативным ядрам, регулируя их функции. Главной из них является интегративная функция.
Подушка получает главные входы от коленчатых тел и неспецифических ядер таламуса. Эфферентные пути от нее идут в височно-теменно-затылочные зоны коры, участвующие в гностических (узнавание объектов и явлений), речевых и зрительных функциях.
Медиодорсалъное ядро получает входы от гипоталамуса, миндалины, гиппокампа, таламических ядер, центрального серого вещества ствола. Проекция этого ядра распр. на ассоциативную, лобную и лимбическую кору. Оно участвует в формировании эмоциональной и поведенческой двигательной активности, а также в запоминании. Разрушение этих ядер устраняет у больных страх, тревогу, напряженность, страдание от боли, но возникает лобный синдром: снижение инициативы, безразличие, гипокинезия.
Латеральные ядра получают зрительную и слуховую импульсацию от коленчатых тел и соматосенсорную импульсацию от вентрального ядра. Интегрированная сенсорная информация от этих источников далее проецируется в ассоциативную теменную кору и используется в функциях гнозиса, праксиса, формировании схемы тела.
3) Неспецифические ядра таламуса — эволюционно более древняя часть, включающая парные интратраламинарную ядерную группу и ретикулярное ядро. Эти ядра содержат нейроны «ретикулярного типа» и функционально рассматриваются как продолжение ретикулярной формации на уровне промежуточного мозга.
Неспецифические ядра имеют многочисленные входы как от других ядер таламуса, так и внеталамические — эти пути проводят преимущественно болевую и температурную чувствительность. В неспецифические ядра поступает непосредственно или через ретикулярную формацию также часть импульсации по коллатералям от всех специфических сенсорных систем. Кроме того, в неспецифические ядра поступает импульсация из моторных центров ствола (красное ядро, черное вещество), ядер мозжечка, от базальных ядер и гиппокампа, а также от коры большого мозга, особенно лобных долей.
При раздражении неспецифических ядер таламуса в ЭЭГ появляется гамма-ритм.
Неспецифические ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ядрами, с другой стороны, объединяя их в единый функциональный комплекс. На кору большого мозга неспецифический таламус оказывает модулирующее (изменяющее состояние) влияние. Разрушение неспецифических ядер не вызывает грубых расстройств эмоций, восприятия, сна и бодрствования, образования условных рефлексов, но нарушает тонкую регулировку поведения. Главная ф-я - модулирующее влияние, обеспечивающее «плавную настройку» ВНД.





