Уксусную, молочную и лимонную кислоты производят также с помощью микроорганизмов. Молочную кислоту получают способом брожения из сахарсодержащего сырья - патоки, крахмала, молочной сыворотки и др.
Молочнокислые бактерии выращивают на средах, содержащих до 15 % сахара. Выход молочной кислоты достигает 60-70 % массы содержащегося в заторе сахара.
Промышленное получение уксуса для пищевых целей основано на уксуснокислом брожении. Уксуснокислые бактерии в специальных чанах на буковых стружках окисляют поступающую питательную среду - уксусно-спиртовой раствор - до уксусной кислоты.
Лимонную кислоту раньше получали из плодов цитрусовых. В настоящее время ее также получают путем брожения. Возбудителем брожения является гриб Аспергиллус нигер, основное сырье - черная патока. Брожение происходит в растворе с содержанием 15 % сахара в аэробных условиях при температуре около 30 °С. Лимонная кислота используется в кондитерской промышленности, производстве безалкогольных напитков, сиропов, кулинарии и медицине.
4. Потребности прокариот в питательных веществах.
Потребность прокариот в питательных веществах
Мономеры, необходимые для построения основных клеточных компонентов, могут быть синтезированы клеткой или поступать в готовом виде из среды. Чем больше готовых соединений должен получать организм извне, тем ниже уровень его биосинтетических способностей, так как химическая организация всех свободноживущих форм одинакова.
Источники углерода
В конструктивном метаболизме основная роль принадлежит углероду, поскольку все соединения, из которых построены живые организмы, — это соединения углерода. Их известно около миллиона. Прокариоты способны воздействовать на любое известное углеродное соединение, т. е. использовать его в своем метаболизме. В зависимости от источника углерода для конструктивного метаболизма все прокариоты делятся на две группы: автотрофы, к которым принадлежат организмы, способные синтезировать все компоненты клетки из углекислоты, и гетеротрофы, источником углерода для конструктивного метаболизма которых служат органические соединения. Понятия "авто-" и "гетеротрофия" характеризуют, таким образом, тип конструктивного метаболизма. Если автотрофия — довольно четкое и узкое понятие, то гетеротрофия — понятие весьма широкое и объединяет организмы, резко различающиеся своими потребностями в питательных веществах.
Наибольшая степень гетеротрофности присуща прокариотам, относящимся к облигатным внутриклеточным паразитам, т. е. организмам, которые могут жить только внутри других живых клеток. Паразитический образ жизни привел к редукции некоторых метаболических путей у этих прокариот, что и обусловило полную их зависимость от метаболизма клетки хозяина.
Другие паразитические прокариотные организмы удается выращивать на искусственных средах, но состав таких сред необычайно сложен. Они содержат, как правило, белки или продукты их неглубокого гидролиза (пептиды), полный набор витаминов, фрагменты нуклеиновых кислот и т. д. Для приготовления питательных сред такого состава используют мясные гидролизаты, цельную кровь или ее сыворотку. Формы, способные расти при создании подходящих условий вне клетки хозяина, называют факультативными паразитами.
Следующую крупную группу прокариот составляют так называемые сапрофиты — гетеротрофные организмы, которые непосредственно от других организмов не зависят, но нуждаются в готовых органических соединениях. Они используют продукты жизнедеятельности других организмов или разлагающиеся растительные и животные ткани. К сапрофитам относится большая часть бактерий. Степень требовательности к субстрату у сапрофитов весьма различна. В эту группу входят организмы, которые могут расти только на достаточно сложных субстратах (молоко, трупы животных, гниющие растительные остатки), т. е. им нужны в качестве обязательных элементов питания углеводы, органические формы азота в виде набора аминокислот, пептидов, белков, все или часть витаминов, нуклеотиды или готовые компоненты, необходимые для синтеза последних (азотистые основания, пятиуглеродные сахара). Чтобы удовлетворить потребность этих гетеротрофов в элементах питания, их обычно культивируют на средах, содержащих мясные гидролизаты, автолизаты дрожжей, растительные экстракты, молочную сыворотку.
Есть прокариоты, требующие для роста весьма ограниченное число готовых органических соединений в основном из числа витаминов и аминокислот, которые они не в состоянии синтезировать сами, и наконец, гетеротрофы, нуждающиеся только в одном органическом источнике углерода. Им может быть какой-либо сахар, спирт, кислота или другое углеродсодержащее соединение. Описаны бактерии из рода Pseudomonas, способные использовать в качестве единственного источника углерода и энергии любое из 200 различных органических соединений, и бактерии, для которых источником углерода и энергии может служить узкий круг довольно экзотических органических веществ. Например, Bacillus fastidiosus может использовать только мочевую кислоту и продукты ее деградации, а некоторые представители рода Clostridium растут только в среде, содержащей пурины. Использовать другие органические субстраты для роста они не могут. Биосинтетические способности этих организмов развиты в такой степени, что они сами могут синтезировать все необходимые им углеродные соединения.
Особую группу гетеротрофных прокариот, обитающих в водоемах, составляют олиготрофные бактерии, способные расти при низких концентрациях в среде органических веществ. Организмы, предпочитающие высокие концентрации питательных веществ, относят к копиотрофам. Если у типичных копиотрофов оптимальные условия для роста создаются при содержании в среде питательных веществ в количестве примерно 10 г/л, то для олиготрофных организмов — в пределах 1—15мг углерода/л. В средах с более высоким содержанием органических веществ такие бактерии, как правило, расти не могут и погибают.
Азот
Азот является одним из четырех основных элементов, участвующих в построении клетки. В расчете на сухие вещества его содержится приблизительно 10%. Природный азот бывает в окисленной, восстановленной и молекулярной формах. Подавляющее большинство прокариот усваивают азот в восстановленной форме. Это соли аммония, мочевины, органические соединения (аминокислоты или пептиды). Окисленные формы азота, главным образом нитраты, также могут потребляться многими прокариотами. Так как азот в конструктивном клеточном метаболизме используется в форме аммиака, нитраты перед включением в органические соединения должны быть восстановлены.
Восстановление нитратов до аммиака осуществляется посредством последовательного действия двух ферментов — нитрат- и нитритредуктазы.
Давно была обнаружена способность отдельных представителей прокариотного мира использовать молекулярный азот атмосферы. В последнее время показано, что этим свойством обладают многие прокариоты, принадлежащие к разным группам: эу- и архебактерии, аэробы и анаэробы, фототрофы и хемотрофы, свободноживущие и симбиотические формы. Фиксация молекулярного азота также приводит к восстановлению его до аммиака.
5. Споры бактерий, их строение и свойства.
При наступлении для бактерий неблагоприятных условий они способны образовывать споры. Неблагоприятными условиями могут быть отсутствие питательных веществ в среде, изменение ее кислотности, высокие или низкие температуры, пересыхание среды и другое.
Образование спор бактериями — это в первую очередь способ пережить неблагоприятные условия среды. В отличие от других организмов у бактерий спорообразование почти не используется для размножения.
Споры бактерий сохраняют жизнеспособность в весьма неблагоприятных условиях внешней среды. Они способны переживать крайне высокие и низкие температуры, сохранять жизнеспособность на протяжении очень многих лет. Так известны бактерии, споры которых могут прорастать через 1000 лет. У других бактерий споры выдерживают кипячение. Бывает, что споры способны пережить температуру меньше -200 градусов Цельсия.
В те времена, когда жизнь на Земле только появилась, и на ней существовали преимущественно только бактерии, возможно погодные условия могли быстро меняться, становиться весьма суровыми. Чтобы выжить, бактерии эволюционно выработали в себе способность к спорообразованию. На сегодняшний день бактерии могут жить там, где другие организмы выжить не могут.
В спорах бактерий все жизненные процессы почти прекращены, цитоплазмы мало, и она густая. Спора покрыта толстой оболочкой, защищающей ее от разрушающих факторов внешней среды. Однако спора содержит все необходимое (в том числе ДНК бактерии), чтобы в благоприятных условиях прорасти и образовать полноценную бактериальную клетку.
Большинство бактерий образуют споры, которые называют эндоспорами. В основном их образуют палочковидные бактерии. «Эндо» значит «внутри». То есть у большинства бактерий споры образуются внутри клетки. При образовании спор происходит впячивание клеточной мембраны, и внутри бактерии обособляется область — будущая спора. Туда переходит ДНК. Вокруг этой области образуется толстый слой так называемой коры, которая будет защищать спору. С ее внутренней и внешней стороны присутствует мембрана. С внешней стороны от мембраны есть еще несколько оболочек.
У палочковидных бактерий эндоспоры могут образовываться в разных местах клетки. У одних — в середине, у других — ближе к концу, у третьих — у самого края палочки-клетки.
Зеленым цветом обозначена оболочка споры, пространство внутри - цитоплазма
Существуют виды бактерий, которые образуют не эндоспоры, а экзоспоры, цисты и другие формы покоящихся форм. «Экзо» говорит о том, что спора образуется не внутри клетки бактерии, а как бы снаружи от нее. Образование экзоспор происходит путем образования своеобразных почек у клетки. После чего такие почки покрываются толстой оболочкой, превращаются в споры и отделяются.
С помощью спор бактерии не только переживают неблагоприятные условия, но и расселяются, так как споры очень легкие и легко разносятся ветром и водой.
Розовым цветом обозначены бактерии, серо-зеленым - их споры. Видно, что споры меньше бактерий
6. Способы размножения бактерий.
Размножение микроорганизмов -- бинарное деление одноклеточных микроорганизмов (бактерий, риккетсий, простейших, дрожжей), в результате которого образуются две новые дочерние полноценные особи, наделенные генетической информацией материнской клетки. Дрожжеподобные грибы могут размножаться почкованием, спорами; плесневые грибы и актиномицеты размножаются обычно спорами.
- Бактерии. Размножаются простым поперечным делением. Бактерии являются гаплоидными клетками. В состав бактериальной клетки входит капсула, клеточная стенка, цитоплазматическая мембрана, цитоплазма, где располагаются мезосомы, рибосомы, нуклеоид, и включения. Некоторые бактериальные клетки имеют жгутики и образуют споры. В отличие от животных клеток такие внутренние структуры бактериальной клетки, как мезосомы, рибосомы, нуклеоид, не имеют мембран, отграничивающих их от цитоплазмы. По способу питания бактерий делят на автотрофов и гетеротрофов, по способу дыхания -- на аэробов и анаэробов.
Актиномицеты. Размножаются спорами и поперечным делением (отшнуровыванием) гиф. Занимают промежуточное положение между грибами и бактериями. Среди лучистых грибов сеть патогенные виды -- возбудители актиномикозов. Многие актиномицеты являются продуцентами антибиотиков. (см. Антибиотики). В «Определителе» Берджи актиномицеты названы стрептомицетами.
Дрожжи. Существует 2 вида размножения дрожжей -- вегетативное (бесполое) и половое с образованием спор. У большинства видов дрожжей вегетативное размножение осуществляется почкованием, редко делением (Schizosaccharomyces). Аспорогенные. дрожжи размножаются только почкованием. Половое размножение происходит при неблагоприятных условиях, когда дрожжи перестают почковаться и превращаются в сумки (аски) со спорами -- аскоспоры. Половой процесс заключается в копуляции (слиянии) 2 вегетативных клеток путем сближения их и образования копуляционного канала, в котором происходит слияние частей плазмы и ядра клеток, называемое кариогамией, с образованием диплоидной зиготы, представляющей 2 клетки, соединенные копуляционным каналом. Редукционное деление, или мейоз, сопровождаемое уменьшением числа хромосом вдвое, происходит сразу, без полового процесса, и зигота превращается в аск с 4 гаплоидными спорами, поэтому вегетативное поколение таких спор гаплоидно. Споры прорастают без копуляции. Так происходит размножение у дрожжей Zygosaccharomyces (рис. 2Б). У дрожжей Saccharomyces половой процесс происходит при слиянии спор или проросших из них клеток с образованием диплоидной зиготы, которая сразу начинает почковаться, образуя диплоидное потомство (рис. 2А). Мейоз происходит непосредственно перед образованием спор.
Плесневые грибы. У Грибов различают вегетативное, половое и бесполое размножение. Вегетативное размножение может осуществляться при отделении от основной массы мицелия его частей, которые могут развиваться самостоятельно, а так же путем почкования мицелия или отдельных клеток у дрожжевых грибов. Половое размножение состоит в слиянии половых клеток, в результате чего возникает зигота. Бесполое размножение осуществляется при помощи специальных образований, называемых спорами. Споры могут развиваться внутри специальных споровместилищ или на концах особых выростов мицелия - конидиеносцах. Основной способ размножения плесневых грибов - при помощи спор. Плесень размножается невероятно быстро.
В обыкновенной хлебной плесени можно различить маленькие чёрные точки - спорангии, в которых образуются споры. В одном спорангии содержится до 50.000 спор, каждая из которых способна воспроизвести сотни миллионов новых спор всего за несколько дней! А если условия благоприятные, плесень быстро появится на книге, обуви или на упавшем дереве в лесу.
Метанобразующие бактерии
Предположение о биологической природе образования метана было высказано еще в XIX в. Однако изучение этого процесса и организмов, его осуществляющих, тормозилось из-за отсутствия чистых культур. Сложность заключается в чрезвычайной чувствительности большинства метанобразующих бактерий к О2. Быстрый прогресс в изучении этой группы архебактерий связан с использованием методов культивирования анаэробов, разработанных Р.Е.Хангейтом (R.E.Hungate). В качестве основных приемов используется удаление О2 из газов, в атмосфере которых осуществляются культивирование и все необходимые для работы операции, а также применение предварительно восстановленных сред.
Метанобразующие бактерии (метаногены) - морфологически разнообразная группа, объединяемая двумя общими для всех ее представителей признаками: облигатным анаэробиозом и способностью образовывать метан. Для создания таксономической структуры метанобразующих бактерий был использован филогенетический подход, основанный на сравнительном анализе нуклеотидных последовательностей 16S рРНК. В соответствии с таким подходом в девятом издании Определителя бактерий Берги группа разделена на три порядка (Methanobacteriales, Methanococcales, Methanomicrobiales), коэффициент сходства (SAB) для которых составляет 0,2-0,28. Далее порядки разделены на 6 семейств (SAB=0,34-0,36) и 13 родов (Sab=0,46-0,51). Число видов достигает более 40. SAB для них колеблется в пределах 0,55-0,65. О гетерогенности группы можно судить и по нуклеотидному составу ДНК ее представителей (молярное содержание ГЦ-оснований - от 27 до 61%).
В состав группы входят бактерии с разной морфологией: прямые или изогнутые палочки разной длины; клетки неправильной формы, близкие к коккам; извитые формы. У некоторых видов наблюдается тенденция формировать нити или пакеты. Клетки неподвижные или передвигающиеся с помощью перитрихиально или полярно расположенных жгутиков. У представителей рода Methanosarcina в клетках найдены газовые вакуоли. Для некоторых метаногенов характерна развитая система внутриклеточных элементарных мембран, являющихся результатом разрастания и впячивания в цитоплазму ЦПМ и сохраняющих с ней связь. У этой группы архебактерий обнаружены клеточные стенки трех типов: состоящие из псевдомуреина, построенные из белковых глобул и гетерополисахаридной природы. Описан микоплазмоподобный метаноген, выделенный в род Methanoplasma, не имеющий клеточной стенки и фильтрующийся через мембранные фильтры с диаметром пор 0,45 мкм.
20-30% мембранных липидов метаногенов представлены нейтральными и 70-80% - полярными липидами. Последние - это в основном два типа простых эфиров глицерина и терпеноидных спиртов (С20-фитаниловый и С40- бифитаниловый), на основе которых образуются полярные фосфо- и гликолипиды (рис. 14 и рис. 102). В зависимости от вида клеточные мембраны могут содержать оба типа эфиров или только один. Основными нейтральными липидами являются С20-, С25- и С30-ациклические изопреноидные углеводороды, насыщенные или содержащие двойные связи. Запасных продуктов в виде поли-бета-оксимасляной кислоты или гликогена в клетках не обнаружено.
Метанобразующие бактерии - строгие анаэробы. Первые исследования чистых культур, выделенных из рубца жвачных животных, показали, что рост их возможен при начальном окислительно-восстановительном потенциале среды ниже -300 мВ. Рост некоторых видов полностью подавляется при содержании в газовой фазе более 0,004% молекулярного кислорода. Однако описаны виды с относительно низкой чувствительностью к О2. В их клетках найдена супероксиддисмутаза. Возможно, в природе такие виды могут сохранять жизнеспособность при кратковременных контактах с О2 и возобновлять рост в анаэробных условиях.
Большинство метанобразующих бактерий имеют температурный оптимум для роста в области 30-40 градусов по С, т.е. являются мезофилами, но есть виды, у которых оптимальная зона сдвинута в сторону более низких (25 градусов по С) или высоких (55-65 градусов по С) температур. Выделен экстремально термофильный организм Methanothermus fervidus, растущий при 55-97 градусов по С (оптимум 80 градусов по С). Все известные представители этой группы - нейтрофилы с оптимальным рН в области 6,5-7,5. Среди метаногенов есть галофилы, требующие в качестве одного из оптимальных условий для роста содержания в среде до 65-70 г/л NaCl.
В качестве источника углерода и энергии для роста метаногены используют узкий круг соединений. Наиболее универсальными источниками углерода и энергии для них является газовая смесь Н2 и СО2. Более 3/4 известных видов утилизируют Н2+СО2. Некоторые метаногены приспособились к облигатному использованию этих соединений. Следующими по распространенности источниками углерода и энергии служат формиат, ацетат, метанол, метиламины и моноокись углерода.
Около половины изученных видов не нуждаются в каких-либо органических соединениях. Для роста многих культур в атмосфере Н2 и СО2 требуется внесение в среду органических веществ, стимулирующих рост или абсолютно для него необходимых. Это могут быть некоторые витамины группы В, ацетат, пируват, сукцинат, отдельные аминокислоты, дрожжевой экстракт или компоненты неизвестного состава, содержащиеся в природных средах обитания. Так, штаммы, выделенные из рубца жвачных животных, нуждаются в добавках рубцовой жидкости. Сложные органические соединения метанобразующие бактерии использовать не могут.
В качестве источника азота метаногены используют аммонийный азот или некоторые аминокислоты. Для ряда видов показана способность к азотфиксации. Источником серы могут служить сульфаты, сульфидили серосодержащие аминокислоты.
9. ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ метанообразующиХ бактерии
Основными местами обитания метанообразующих бактерий являются осадки пресных, морских, гиперсоленых водоемов, гидротермы, очистные сооружения, заболоченные почвы. Кроме того, эти бактерии являются важными компонентами пищеварительного тракта растительноядных животных; найдены они в толстом кишечнике человека (таблица 2).
Субстратная специфичность метанообразующих бактерий обусловливает необходимость их взаимодействия с другими анаэробными бактериями - продуцентами необходимых метаногенам субстратов. В этой связи метаногены в природе развиваются в составе сложного общества, которое обеспечивает в анаэробных условиях разложение практически любого органического соединения. Конечной стадией такого разрушения является образование метана, который, насколько известно, не может быть использован в анаэробных реакциях.
В основе взаимодействия микроорганизмов, входящих в сообщество, лежит трофическая цепь, ведущая от сложных биополимеров к простым газообразным продуктам. При этом на первом этапе, благодаря жизнедеятельности "первичных анаэробов", идет деструкция высоко-энергизованных природных соединений с образованием летучих жирных кислот (формиата, ацетата, пропионата, бутирата), органических кислот (лактата, сукцината), низших спиртов (этанола, метанола), альдегидов, кетонов, водорода и углекислого газа. На втором этапе действуют "вторичные анаэробы", которые используют перечисленные выше продукты в качестве энергетических субстратов. В результате образуется ацетат, Н2, СО2, формиат. В последнюю очередь происходит образование метаногенами метана, который далее удаляется из системы в анаэробную зону в виде пузырьков газа, где используется метанотрофными бактериями.
Согласно современным представлениям процесс образования метана происходит по двум механизмам. Первый механизм начинается с последовательности процессов, приводящих к центроболитам (НСООН, СН3ОН и др.), которые затем используются метанообразующими бактериями. Этот механизм метаногенеза связан с переносом углеродсодержащего материала в трофической цепи.
Второй механизм обусловлен потреблением метаногенами Н2. Следствием этого процесса является изменение метаболизма первичных анаэробов в сторону увеличения выхода энергии. Тесное взаимодействие между микроорганизмами на основе межвидового переноса водорода получило название синтрофии.
Метанообразующие бактерии представляют определенный практический интерес как продуценты газообразного топлива – метана. В институте биохимии им. Баха разработан способ получения в процессе метанового брожения кормового витамина В12 при сбраживании отходов, богатых углеводами.
Метанообразующие бактерии являются одним из основных компонентов активного ила метантенков – анаэробных очистных сооружений сточных вод. В данном случае бактерии используются для сбраживания углеродсодержащих осадков. Многие продукты анаэробного распада в метантенках могут быть утилизированы. Выделяющийся в процессе метанового брожения метан собирается в газгольдеры и используется на отопление очистной станции, в том числе и на поддержание требуемой температуры в метантенке. Сброженный осадок после подсушивания на иловых площадках или механического обезвоживания может быть использован в качестве удобрения на сельскохозяйственных полях.
10 Дыхание микроорганизмов, сущность процесса. Способы дыхания.
Для своей жизнедеятельности каждая микробная клетка кроме питательных веществ нуждается в энергии. Эту энергию микроорганизмы получают в процессе дыхания. Сущность дыхания у микробов заключается в окислении сложных органических соединений до более простых веществ с выделением тепловой энергии, которая и используется микробами. В большинстве случаев микроорганизмы получают энергию путем окисления углеводов и других органических соединений. За счет полученной энергии происходит синтез сложных органических соединений в самой клетке.
По типу дыхания микроорганизмы делятся на аэробы и анаэробы. Аэробы - микроорганизмы, которые для дыхания и получения необходимой энергии нуждаются в свободном доступе кислорода из воздуха. У этой группы микробов процесс дыхания аэробный. Анаэробы - микроорганизмы, которые получают энергию при дыхании без доступа кислорода воздуха путем расщепления питательных веществ. Различают облигатные (строгие) и факультативные (нестрогие) анаэробы. Облигатные анаэробы проявляют свою жизнедеятельность только при отсутствии кислорода воздуха. Факультативные анаэробы могут развиваться в средах как в присутствии кислорода воздуха, так и без него.
В химизме дыхательных процессов у аэробов и анаэробов имеется много общего. Во всех случаях первым этапом дыхательных процессов является отщепление водорода от субстрата (дегидрирование) в присутствии специфических ферментов - дегидрогеназ. Происходящие процессы носят окислительно-восстановительный характер.
Сущность окисления состоит в потере электронов окисляющимся веществом, тогда как сущность восстановления состоит в присоединении этих электронов восстанавливающимся веществом. Та или иная последовательность биохимических реакций в течение обменных процессов возможна благодаря тонким изменениям окислительно-восстановительного потенциала, под которым понимают способность вещества отдавать или получать электроны.
При аэробном типе дыхания аэробные дегидрогеназы передают отнятый от субстрата водород или непосредственно кислороду воздуха, или цитохромной системе. Это так называемое аэробное дегидрирование, при котором происходит обычно полное окисление. При полном окислении конечными продуктами являются вода и углекислота, при этом освобождается вся энергия. При неполном окислении происходит образование продуктов, в которых заключается значительная часть энергии.
В анаэробных условиях биохимические процессы происходят при отсутствии кислорода воздуха. Анаэробные дегидрогеназы не могут отдавать водород кислороду воздуха, а передают его другим веществам, от которых сравнительно легко отщепляется кислород. Это так называемое анаэробное дегидрирование, при котором происходит неполное окисление субстрата.
Дыхание микроорганизмов представляет собой биологическое окисление различных органических соединœений и некоторых минœеральных веществ. В итоге окислительно-восстановительных процессов и брожения образуется тепловая энергия, часть которой используется микробной клеткой, а остальное количество выделяется в окружающую среду. Сегодня окисление определяют как процесс отнятия водорода (дегидрирование), а восстановление — его присоединœения. Эти термины применяют к реакциям, связанным с переносом протонов и электронов или только электронов. При окислении вещества происходит потеря электронов, а при восстановлении — их присоединœение. Считают, что перенос водорода и перенос электронов — эквивалентные процессы.
Энергия, освобождаемая в процессе окислительно-восстановительных реакций, накапливается в макроэргических соединœениях АДФ и АТФ (аденозиндифосфат и аденозинтрифосфат). Эти соединœенияимеют макроэргические связи, обладающие большим запасом биологически доступной энергии. Οʜᴎ локализованы в сложно устроенных структурах микробных клеток — мезосомах, или митохондриях.
По типу дыхания микроорганизмы делят на аэробов, анаэробов и факультативных анаэробов.
Аэробное дыхание микроорганизмов — это процесс, при котором последним акцептором водорода (протонов и электронов) является молекулярный кислород. В результате окисления главным образом сложных органических соединœений образуется энергия, которая выделяется в среду или накапливается в макроэргических фосфатных связях АТФ. Различают полное и неполное окисление.
Полное окисление. Основной источник энергии у микроорганизмов —углеводы. В результате расщепления глюкозы в аэробных условиях процесс окисления идет до образования диоксида углерода и воды с выделœением большого количества свободной энергии:
С6Н12О6 + 6О2 → 6СО2 + 6Н2О + 674 ккал.
Неполное окисление. Не всœе аэробы доводят реакции окисления до конца. При избытке углеводов в среде образуются продукты неполного окисления, в которых заключена энергия. Конечными продуктами неполного аэробного окисления сахара бывают органические кислоты: лимонная, яблочная, щавелœевая, янтарная и другие, которые образуются плесневыми грибами. Так же осуществляется аэробное дыхание уксуснокислыми бактериями, в которых при окислении этилового спирта образуется не диоксид углерода и вода, а уксусная кислота и вода:
С2Н5ОН + О2 → СН3СООН + Н2О + 116 ккал.
этиловый спирт уксус. к-та
Окисление этилового спирта уксуснокислыми бактериями может идти и дальше — до появления диоксида углерода и воды, при этом освобождается большое количество энергии:
С2Н5ОН + 3О2 → 2СО2 + 3Н2О + 326 ккал.
этиловый спирт
Анаэробное дыхание осуществляется без участия молекулярного кислорода. Различают собственно анаэробное дыхание (нитратное, сульфатное) и брожение. При анаэробном дыхании акцептором водорода являются окисленные неорганические соединœения, которые легко отдают кислород и превращаются в более восстановленные формы. Нитратное дыхание — восстановление нитратов до молекулярного азота. Сульфатное дыхание — восстановление сульфатов до сероводорода.
Брожение — расщепление органических углеродсодержащих соединœений в анаэробных условиях. Оно характеризуется тем, что последнимакцептором водорода служит молекула органического вещества с ненасыщенными связями. Вещество при этом разлагаетсятолько до промежуточных продуктов, представляющих собой сложныеорганические соединœения (спирты, органические кислоты). Заключенная в них энергия не используется микробами, а об  разовавшаяся в небольших количествах энергия выделяется в окружающую среду.
разовавшаяся в небольших количествах энергия выделяется в окружающую среду.
Типичными примерами анаэробного дыхания являются:
- спиртовое брожение (дыхание дрожжей в анаэробных условиях):
С6Н12О6 → 2С2Н5ОН + 2СО2 + 27 ккал;
этилов. спирт
- молочнокислое брожение (дыхание молочнокислых бактерий):
С6Н12О6 → 2С3Н6О3 + 18 ккал;
молочн. к-та
- маслянокислое брожение (дыхание маслянокислых бактерий):
С6Н12О6 → С3Н7СООН + 2СО2 + 2Н2 + 15 ккал;
маслян. к-та
Как видно из приведенных уравнений, при анаэробном дыхании освобождается значительно меньше энергии, чем при аэробном. По этой причине при анаэробном дыхании для того, чтобы обеспечить потребность в крайне важно м количестве энергии, микроорганизмам крайне важно потреблять больше сахаров, чем при аэробном.
Большая часть энергии, образующейся при дыхании, освобождается в окружающую среду. Это вызывает нагревание продуктов, в которых развиваются микроорганизмы. Именно так нагревается вино, в котором происходит спиртовое брожение; нагревается влажное зерно, торф, сено.
13 Уксусную кислоту можно получать следующими способами:
1) Из метана, являющегося основным компонентом природного газа, получают ацетилен:
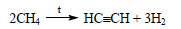
Ацетилен в присутствии сульфата ртути (II) присоединяет воду, образуется уксусный альдегид (реакция Кучерова):
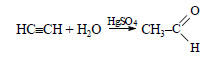
При окислении уксусного альдегида образуется уксусная кислота:
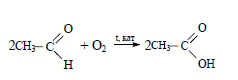
2) При гидратации этилена, являющегося побочным продуктом крекинга нефти, получают этиловый спирт:
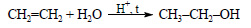
При окислении этилового спирта получают уксусную кислоту:
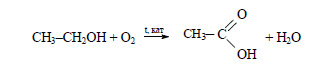
3) Уксусную кислоту можно получить при окислении бутана в присутствии катализатора:
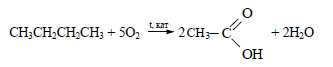
Наиболее перспективным является последний метод, так как бутан является компонентом природного газа и попутных нефтяных газов, а уксусная кислота получается в одну стадию.
Как одна из простейших органических кислот, уксусная кислота является естественным метаболитом (промежуточным продуктом жизнедеятельности) очень многих живых организмов. В естественной среде, например, она является типичным промежуточным звеном концевого окисления этилового спирта CH3-CH2-OH (его формула в ином представлении — C2H5OH), что и определило её первичный способ получения, известный человеку — сбраживание сахаров (прежде всего глюкозы, фруктозы и их производных) напрямую либо через промежуточные стадии спирта этилового и альдегида уксусного (CH3-CHO). В зависимости от происхождения используемого сырья/способа производства, к слову «уксус» нередко добавляется соответствующее прилагательное: «винный уксус», «рисовый уксус», «яблочный уксус» и так далее — и все эти вещества издревле использовались при приготовлении пищи и консервировании/обработке разнообразных пищевых/непищевых продуктов.
Помимо использования в быту и пищевой индустрии (уксусная кислота является официально зарегистрированной пищевой добавкой/консерв
антом с кодом E-260), она также имеет большое промышленное значение — и как компонент моющих/чистящих средств, продуктов гигиены и дезинфекции, и как промежуточное сырьё либо технический продукт (например, специфический растворитель в целом ряде промышленных синтезов). Поскольку массовое производство технической уксусной кислоты «естественным» путём микробиологической переработки пищевых продуктов дорого/неэффективно и имеет смысл лишь в случае наличия некоторых типов бросового либо условно-бесплатного сырья, постоянно изыскивались способы иного её получения.
14 Образование лимонной кислоты
Лимонная кислота содержится в различных растениях. Она была впервые выделена К. В. Шееле из лимонного сока, где её содержание достигает 10%. В промышленности лимонную кислоту получают путем лимоннокислого брожения отходов сахарного производства с помощью плесневого грибка Aspergillius niger
Применение лимонной кислоты
Лимонная кислота нашла широкое применение в пищевой промышленности (производство плавленых сыров, напитков и т.д.), при производстве косметики, цемента, а также в нефтехиии.
Соли лимонной кислоты называются цитратами. Цитрат натрия применяется для консервации донорской крови. Противосвертывающее действие основано на том, что цитрат натрия связывает участвующие в процессе свертывания ионы кальция в нерастворимый цитрат кальция.
15 Молочную кислоту открыл шведский химик Карл Шееле. В 1780 году он выделил её из прокисшего молока в виде коричневого сиропа. Французский химик Анри Браконно обнаружил, что она образуется при молочнокислом брожении[1].
В 1807 году Йенс Якоб Берцелиус выделил из мышц цинковую соль молочной кислоты.
Молочная кислота – это органическое соединение с формулой CH3CH(OH)CO2H. Визуально это белое, твердое, растворимое в воде вещество или прозрачная жидкость, которая может быть натуральной или вырабатываться синтетическим путем. Молочная кислота (пищевая добавка Е270) состоит из двух оптических изомеров. Один из них известен как L-(+)-молочная кислота, а другой, его зеркальное отражение, D-(-)-молочная кислота. Их смесь в равных количествах называется DL-молочная кислота.
В 1856 году Луи Пастер обнаружил лактобактерии и выяснил их роль в создании молочной кислоты. Эти бактерии способны также размножаться во рту. Кислота, которую они производят, приводит к заболеванию зубов, которое известно как кариес. В промыш






