Классификация нейронов
I. По строению (количеству отростков) (рис. 2):
1. Униполярные (одноотростчатые) – это клетки, от тела которых отходит только один отросток. В нервной системе человека почти не встречаются.
2. Биполярные (двухотростчатые) – это клетки, которые имеют один аксон и один дендрит. Они характерны для зрительной, слуховой, обонятельной сенсорных систем.
3. Мультиполярные (многоотростчатые) – мелкие мультиполярные нейроны являются ассоциативными, средние и крупные – мультиполярными, пирамидные нейроны – двигательными и эффекторными; имеют один аксон и множество дендритов. К такому типу нейронов принадлежит большинство нейронов ЦНС. Исходя из особенностей формы этих клеток их делят на веретенообразные, корзинчатые, звездчатые, пирамидные.
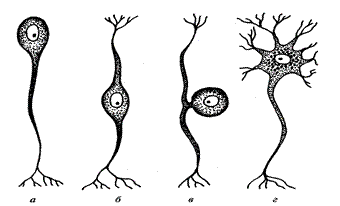
Рис. 2. Нейроны:
а – униполярный нейрон; б – биполярный нейрон;
в – псевдоуниполярный нейрон; г – мультиполярный
4. Псевдоуниполярные (ложноодноотростчатые) – отросток как бы один, но он раздваивается. Такие клетки всегда чувствительные и всегда находятся вне ЦНС в чувствительных ганглиях. Для этих клеток характерна определенная локализация. Они принадлежат неспецифическим сенсорным модальностям (болевая, температурная, тактильная, проприоцептивная).
II. В зависимости от отдела ЦНС различают нейроны соматической и вегетативной нервной системы.
III. По виду медиатора, выделяющегося в окончаниях аксонов, различают нейроны адренергические, холинергические, дофаминергические и т.д.
IV. По влиянию на другую клетку – возбуждающие, тормозные.
V. По специфичности воспринимаемой сенсорной информации нейроны высших отделов ЦНС могут быть мономодальные, бимодальные, полимодальные.
VI. По активности – фоновоактивные и «молчащие». Фоновоактивные нейроны играют важную роль в поддержании тонуса ЦНС. «Молчащие» нейроны активируются только при действии раздражителей.
VI. По функции выделяют три типа нейронов:
1) чувствительные;
2) вставочные;
3) двигательные.
VII. Центральные и периферические (в чувствительных и вегетативных ганглиях).
Тела чувствительных нейронов всегда находятся вне ЦНС, в чувствительных ганглиях – спинномозговых или черепных нервах. Дендриты чувствительных нейронов оканчиваются чувствительными нервными окончаниями – рецепторами.
Двигательные нейроны посылают импульсы к рабочему органу. Двигательный нейрон обеспечивает ответную реакцию на раздражение (например, сокращается мышца).
Вставочные нейроны обеспечивают передачу нервного импульса с чувствительного нейрона на двигательный нейрон. И двигательные, и вставочные нейроны находятся в пределах ЦНС.
Нейроглии
Помимо нейронов в ЦНС имеются глиальные клетки (нейроглии). Термин «глия» означает «связывающее» и отражает роль нейроглии как посредника между кровеносными сосудами и нейронами. В ЦНС человека количество глиальных клеток на порядок больше, чем количество нейронов и составляет около 50% объема ЦНС. Величина мембранного потенциала глиальных клеток больше (-90 мВ), чем у нейрона (-70 мВ), поэтому повышение концентрации ионов К+ во внеклеточной среде приводит к деполяризации глиальной мембраны раньше, чем нервной. Глиальная мембрана является активным калиевым электродом. Она не способна генерировать потенциал действия, поэтому возникают только локальные потенциалы. Между клетками глии существуют участки низкоомной связи, которые обеспечивают обмен ионов и мелких молекул. Клетки нейроглии заполняют промежутки между нервными клетками. В мозге нет соединительной ткани, вместо нее – нейроглия. Периферические аксоны (находящиеся за пределами ЦНС) также окружены оболочкой из глиальных клеток. Они способны к делению в течение всей жизни. С возрастом количество нейронов уменьшается, а нейроглий – увеличивается.
Существует несколько типов нейроглии. В ЦНС они представлены астроцитами и олигодендроцитами, а в периферической нервной системе – шванновскими клетками и ганглионарными глиоцитами. Кроме того, центральными глиальными клетками считаются клетки микроглии и клетки эпендимы.
Функции клеток глии:
1) опорная – в цитоплазме астроцитов находятся глиальные филаменты, выполняющие в ткани ЦНС механическую опорную функцию;
2) защитная (цитокинез, фагоцитоз) – в случае повреждения отростки астроцитов, содержащие глиальные филаменты, подвергаются гипертрофии и формируют глиальный рубец. При повреждении мозговой ткани микроглия наряду с проникающими в ЦНС из кровотока фагоцитами способствует удалению продуктов клеточного распада;
3) являются трофическим аппаратом;
4) регулируют определенную концентрацию ионов кальция и калия в межклеточном пространстве;
5) активно поглощают нейромедиаторы;
6) обеспечивают электрическую изоляцию аксонов путем формирования миелиновой оболочки.
Глиальные клетки выполняют в нервной системе множество еще не совсем изученных функций.
Классификация рецепторов
I. По локализации выделяют следующие виды рецепторов:
1) экстерорецепторы – располагаются в коже и слизистых оболочках и воспринимают внешнее раздражение (терморецепторы, рецепторы осязания (тактильные), давления, болевые). К ним также относятся чувствительные элементы органов чувств, например, палочки и колбочки сетчатки;
2) рецепторы, заложенные в аппарате движения (в мышцах, сухожилиях, суставных капсулах и связках, надкостнице, фасциях), называются проприорецепторами (глубокими рецепторами). Они воспринимают чувство веса, давления, вибрации, состояние мышц;
3) интерорецепторы – находятся в стенках внутренних органов и сосудов. Воспринимают химический состав определенных веществ, степень наполнения внутренних органов;
4) рецепторы специализированных органов чувств. Отвечают за восприятие зрительных, слуховых, обонятельных, вкусовых раздражений и чувство равновесия.
II. По строению выделяют следующие виды рецепторов:
1) свободные нервные окончания: воспринимают болевые импульсы;
2) инкапсулированные: воспринимают тактильные, температурные и проприорецептивные раздражения;
3) первичночувствующие клетки: воспринимают зрительные, слуховые, вестибулярные и вкусовые раздражения.
III. По природе адекватного раздражителя:
– механорецепторы;
– хеморецепторы;
– фоторецепторы;
– терморецепторы;
– осморецепторы;
– волюморецепторы и т.д.
IV. По характеру ощущений:
– зрительные;
– слуховые;
– вкусовые;
– тактильные;
– температурные;
– болевые;
– положения тела в пространстве.
V. По числу воспринимаемых раздражителей:
– мономодальные;
– полимодальные.
Участок тела, содержащий рецепторы, при раздражении которых возникает определенный рефлекс, называется рецептивным полем рефлекса или рефлексогенной зоной. Любой рефлекс можно вызвать только с определенного рецептивного поля. Например, коленный рефлекс (разгибание колена) возникает при ударе по сухожилию ниже надколенника.
Любое раздражение, воспринимаемое рецептором, кодируется (преобразуется) в нервный импульс и направляется по афферентным волокнам в ЦНС. В центре информация перерабатывается, отбирается, передается на двигательные волокна, которые посылают возбуждение к рабочим органам. В результате вызывают тот или иной приспособительный акт – движение или секрецию.
Во время ответной реакции возбуждаются рецепторы рабочего органа и от них в ЦНС поступают импульсы, т.е. информация о достигнутом результате. Афферентные импульсы осуществляют обратную связь, либо усиливают и уточняют реакцию, если она не достигла цели, либо прекращают ее, т.е. работают по принципу обратной связи.
Для осуществления любого рефлекса необходима целостность всех звеньев рефлекторной дуги. Нарушение хотя бы одного из них ведет к исчезновению рефлекса. Если лапку лягушки опустить в слабый раствор серной кислоты, возникнет оборонительный рефлекс – лапка отдернется. Однако если снять кожу и тем самым удалить кожные рецепторы, то серная кислота не окажет воздействия.
Подобное можно наблюдать при разрушении любого другого звена: ЦНС, чувствительных или двигательных нервных волокон. Самое сильное раздражение не вызовет ответной реакции, нервная деятельность будет отсутствовать.
Время рефлекса. Время, прошедшее от момента нанесения раздражения до ответа на него, называется временем рефлекса. Это время слагается из времени, необходимого для возбуждения рецепторов, проведения возбуждения по чувствительным волокнам ЦНС, двигательным волокнам и периода возбуждения рабочего органа. На проведение возбуждения через нервные центры уходит большая часть времени – центральное время рефлекса. Возбуждение проводится через синапсы, а в синапсах ЦНС происходит замедление проведения возбуждения. Чем меньше нейронов входит в состав рефлекторной дуги, тем короче время рефлекса. Простые сухожильные рефлексы, имеющие моносинаптическую (двухнейронную) рефлекторную дугу, наиболее быстрые, так как возбуждение проходит через один синапс. Время сложных вегетативных рефлексов дольше. Время рефлекса зависит от силы раздражения и возбудимости ЦНС. При сильном раздражении оно короче, при снижении возбудимости время рефлекса увеличивается, при повышении возбудимости – уменьшается.
Классификация рефлексов
I. По характеру связи между нейронами различают:
1. Безусловные (врожденные) рефлексы. Они передаются по наследству, присущи каждому биологическому виду. Их дуги формируются к моменту рождения и в норме сохраняются в течение всей жизни. Однако они могут изменяться под влиянием болезни.
2. Условные (рефлекторные реакции, приобретаемые на протяжении индивидуальной жизни человека, например, протягивание руки к выключателю). Они возникают при индивидуальном развитии и накоплении новых навыков. Выработка новых временных связей зависит от изменяющихся условий среды. Условные рефлексы формируются на основе безусловных и с участием высших отделов головного мозга.
II. По расположению рецепторов, раздражение которых вызывает данный рефлекторный акт:
1) экстерорецептивный рефлекс – раздражение рецепторов внешней поверхности тела (зрительные, обонятельные и др.);
2) висцеро- или интерорецептивный рефлекс – возникающий при раздражении рецепторов внутренних органов и сосудов (хемо-, баро-, осмо-);
3) проприорецептивный (миотатический) рефлекс – раздражение рецепторов скелетных мышц, суставов, сухожилий.
III. По биологическому значению:
1. Пищевые:
– рефлекторный акт глотания;
– жевания;
– сосания;
– слюноотделения;
– секреции желудочного и поджелудочного сока и др.
2. Оборонительные – реакции устранения от повреждающих и болевых раздражений.
3. Рефлексы сохранения гомеостаза:
– рефлекс терморегуляции;
– дыхательный рефлекс;
– сердечный;
– сосудистые, способствующие сохранению постоянства артериального давления и др.
4. Позно-тонические (рефлексы положения тела в пространстве).
5. Локомоторные (рефлексы передвижения тела в пространстве).
6. Половые – рефлексы, связанные с осуществлением полового акта; в эту же группу можно отнести и так называемые родительские рефлексы, связанные с выкармливанием и выхаживанием потомства.
7. Стато-кинетические и локомоторные – рефлекторные реакции поддержания определенного положения и передвижения тела в пространстве.
8. Ориентировочный рефлекс – рефлекс на новизну. Он возникает в ответ на любое достаточно быстро происходящее колебание окружающей среды и выражается внешне в настораживании, прислушивании к новому звуку, обнюхивании, повороте глаз и головы, а иногда и всего тела в сторону появившегося светового раздражителя и т.п. Осуществление этого рефлекса обеспечивает лучшее восприятие действующего агента и имеет важное приспособительное значение.
IV. По эффекторам:
1. Соматические (скелетная мускулатура).
2. Вегетативные (гладкая мускулатура или железы), т.е. эффектором являются внутренние органы, эфферентная часть сформирована вегетативными нейронами.
V. По уровню замыкания рефлекторной дуги в ЦНС выделяют (рис. 4):
1. Спинальные (рефлекторные дуги замыкаются в спинном мозге). Например, сгибательный рефлекс – укол или нанесение слабого раствора кислоты на лапку лягушки вызывает рефлекторное сокращение мышц этой лапки, которая сгибается и устраняется от раздражителя;
– рефлекс натирания – прикладывание к коже боковой поверхности тела лягушки кусочка фильтровальной бумаги, смоченной кислотой, влечет за собой сокращение приводящих мышц лапки той же стороны, потирание раздраженного места и сбрасывание бумаги;
– рефлекс почесывания – потирание кожи на боку собаки влечет за собой притягивание задней лапы со стороны раздражения к боковой поверхности туловища и ритмические сгибательные движения почесывания;
– коленный рефлекс – при легком коротком ударе по сухожилию четырехглавой мышцы бедра под коленной чашечкой происходит резкое разгибание ноги в колене;
– ахиллов рефлекс – при ударе по ахиллову сухожилию происходит резкое сокращение икроножной мышцы;
– подошвенный рефлекс – раздражение кожи подошвенной части ноги взрослого человека вызывает рефлекторное сгибание стопы и пальцев.
2. Бульбарные (замыкающие в продолговатом мозге).
Например, сосательный рефлекс – прикосновение к губам грудного младенца ведет к появлению ритмических сосательных движений; а также корнеальный рефлекс – прикосновение к роговице глаза ведет к смыканию век.
3. Мезэнцефальные (участвуют нейроны среднего мозга).
Например, зрачковый рефлекс – освещение ярким светом глаза вызывает сужение зрачка
4. Диэнцефальные (нейроны промежуточного мозга).

Рис. 4. Уровни
рефлекторной дуги
5. Корковый (для которого необходимы нейроны коры головного мозга).
VI. По количеству нейронов, участвующих в замыкании рефлекторной дуги:
1) моносинаптические – состоят из двух нейронов (чувствительного и двигательного), импульсы проходят через один синапс;
2) полисинаптические – состоят из трех и более нейронов (чувствительный, двигательный, вставочный), импульсы проходят через несколько синапсов.
VII. По характеру ответной реакции, в зависимости от того, какие органы в ней участвуют:
– моторные, или двигательные рефлексы, – исполнительным органом служат мышцы;
– секреторные рефлексы – заканчиваются секрецией желез;
– сосудодвигательные рефлексы – проявляющиеся в сужении или расширении кровеносных сосудов.
По характеру рефлексов судят о состоянии различных отделов нервной системы. При исследовании рефлексов определяют их уровень, равномерность, асимметрию; при повышенном уровне отмечают рефлексогенную зону. При описании рефлексов применяют следующие градации: живые рефлексы; гипорефлексия; гиперрефлексия (с расширенной рефлексогенной зоной); арефлексия (отсутствие рефлексов).
Сухожильные и надкостничные рефлексы вызываются с помощью короткого удара по сухожилию или надкостнице и имеют важное диагностическое значение в неврологической практике. Рефлекторная реакция проявляется в виде резкого сокращения мышц.
Эта классификация приемлема для простых рефлексов, направленных на объединение функций внутри организма. При сложных рефлексах, в которых участвуют нейроны, находящиеся в высших отделах ЦНС, как правило, в осуществление рефлекторной реакции вовлекаются различные исполнительные органы, в результате чего происходит изменение соотношения организма с внешней средой, а также его поведения.
Нервный центр
С точки зрения анатомии нервный центр (ядро) – совокупность близко расположенных нейронов на определенном уровне мозга, объединенных общими морфологическими особенностями. Например, в продолговатом мозге – центры дыхания, кровообращения и другие.
С точки зрения физиологии нервный центр – это функционально связанная совокупность нейронов, расположенных на различных уровнях ЦНС и обеспечивающих регуляцию той или иной функции или осуществление целостной реакции организма. Например, нервный центр, регулирующий дыхание, имеет свое представительство в разных отделах ЦНС. Например, кора больших полушарий регулирует произвольную регуляцию дыхания; лимбическая кора – связь эмоций и дыхания; гипоталамус – приспособление дыхания к уровню обменных процессов и т.д.
Каждый рефлекс имеет свой центр: существует центр коленного рефлекса, центр локтевого рефлекса, мигательного; есть сердечно-сосудистый, дыхательный, пищевой центры, центры сна и бодрствования, голода и жажды и т.д. В целом организме при формировании сложных адаптивных процессов происходит функциональное объединение нейронов, расположенных на различных уровнях ЦНС, т.е. сложное объединение большого количества центров.
Объединение нервных центров (ядер) между собой осуществляется проводящими путями ЦНС с помощью нейро-нейрональных (межнейронных) синапсов. Существует три типа соединения нейронов: последовательное, дивергентное и конвергентное.
Нервные центры обладают рядом характерных функциональных свойств, которые во многом обусловлены этими тремя типами нейронных сетей, а также свойствами межнейронных синапсов.
Свойства нервных центров
Нервные центры обладают рядом общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Возбуждение в нервных центрах распространяется односторонне – от рецептора к эффектору. Одностороннее проведение связано с тем, что связь между нейронами осуществляется в основном с помощью синапсов, что обусловливается свойством химических синапсов односторонне проводить возбуждение от пресинаптической мембраны к постсинаптической, на которой имеются специфические структуры (рецепторы). Если перерезать передние и задние корешки спинного мозга и раздражать их, то при раздражении задних (чувствительных корешков) с помощью осциллографа можно зарегистрировать возникновение потенциалов на передних корешках, а если раздражать передние, то на задних корешках потенциалы не регистрируются.
2. Центральная задержка. Возбуждение в нервных центрах проводится медленнее, чем по нервному волокну. Это обусловлено замедленным проведением возбуждения через синапсы (синаптическая задержка), которых в ядре много. Синаптическая задержка включает в себя время выделения медиатора из везикул, время трансфузии его через синаптическую щель к постсинаптической мембране и время генерации постсинаптического потенциала.
Центральное время рефлекса – самое короткое у моносинаптического рефлекса, например, время центрального торможения коленного рефлекса – 3 мс. Чем больше нейронов входит в рефлекторную дугу (тем больше синапсов), тем больше время рефлекса.
3. В нервных центрах осуществляется суммация возбуждений. Суммация – сложение допороговых импульсов. Основана на свойстве каждого нейрона в центре к суммации как возбуждения, так и торможения. Различают два вида суммации.
 Временная, или последовательная, если импульсы возбуждения приходят к нейрону по одному и тому же пути через один синапс с интервалом меньшим, чем время полной реполяризации постсинаптической мембраны (рис. 5). В этих условиях локальные токи на постсинаптической мембране воспринимающего нейрона суммируются и доводят ее деполяризацию до уровня Ек, достаточного для генерации нейроном потенциала действия. Известно, что один подпороговый афферентный стимул не вызывает ответной рефлекторной реакции, а создает в ЦНС местное возбуждение. При действии серии подпороговых импульсов выделяется достаточное количество медиатора для того, чтобы деполяризация достигла критического уровня и возникло распространяющееся возбуждение.
Временная, или последовательная, если импульсы возбуждения приходят к нейрону по одному и тому же пути через один синапс с интервалом меньшим, чем время полной реполяризации постсинаптической мембраны (рис. 5). В этих условиях локальные токи на постсинаптической мембране воспринимающего нейрона суммируются и доводят ее деполяризацию до уровня Ек, достаточного для генерации нейроном потенциала действия. Известно, что один подпороговый афферентный стимул не вызывает ответной рефлекторной реакции, а создает в ЦНС местное возбуждение. При действии серии подпороговых импульсов выделяется достаточное количество медиатора для того, чтобы деполяризация достигла критического уровня и возникло распространяющееся возбуждение.
Данная суммация называется временной, потому что на нейрон в течение некоторого промежутка времени приходит серия импульсов (раздражений). Последовательной она называется, потому что реализуется в последовательном соединении нейронов.
Пространственная, или одновременная, – наблюдается в том случае, когда импульсы возбуждения поступают к нейрону одновременно через разные синапсы (рис. 6).
Пространственной данная суммация называется, потому что раздражитель действует на некоторое пространство рецептивного поля, т.е. несколько (минимум два) рецепторов разных участков рецептивного поля. Тогда как временная суммация может реализоваться при действии серии раздражителей на один и тот же рецептор. Одновременной она называется, потому что информации к нейрону приходят одновременно по нескольким (минимум два) каналам связи, т.е. одновременная суммация реализуется конвергентным соединением нейронов.
Рис. 6. Пространственная суммация в результате одновременно
наносимых раздражений:
а – передача возбуждения с одного аксона (уменьшение мембранного потенциала)
б – передача возбуждения с трех аксонов и генерация потенциала действия
Пространственную суммацию, как и последовательную, можно объяснить тем, что при подпороговом раздражении, пришедшем по одному афферентному пути, выделяется недостаточное количество медиатора для того, чтобы вызвать деполяризацию мембраны до критического уровня. Если же импульсы приходят одновременно несколькими афферентным путям к одному и тому же нейрону, в синапсах выделяется достаточное количество медиатора, необходимого для пороговой деполяризации и возникновения потенциала действия.
4. Трансформация ритма возбуждения – изменение количества импульсов возбуждения, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему.
Различают два вида трансформации:
1) понижающая, в основе которой лежит явление суммации возбуждений, когда в ответ на несколько пришедших допороговых возбуждений к нервной клетке в нейроне возникает только одно пороговое возбуждение;
2) повышающая, в ее основе лежат механизмы умножения (мультипликации), способные резко увеличить количество импульсов возбуждения на выходе.
5. Рефлекторное последействие заключается в том, что рефлекторная реакция заканчивается позже прекращения действия раздражителя (рис. 7).
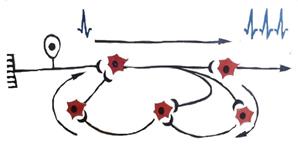
Рис. 7. Кольцевые связи в нервном центре (Лоренто де Но).
Стрелками показано направление движения импульсов
Это явление обусловлено двумя причинами:
1) длительной следовой деполяризацией мембраны нейрона на фоне прихода мощной афферентации (сильной чувствительной импульсации), вызывающей выделение большого количества (квантов) медиатора, что обеспечивает возникновение нескольких потенциалов действия на постсинаптической мембране и, соответственно, кратковременное рефлекторное последействие;
2) пролонгированием выхода возбуждения к эффектору в результате циркуляции (реверберации) возбуждения в нейронной сети типа «нейронной ловушки». Возбуждение, попадая в такую сеть, может длительное время циркулировать в ней, обеспечивая длительное рефлекторное последействие. Возбуждение в такой цепочке может циркулировать до тех пор, пока какое-либо внешнее воздействие затормозит этот процесс или в ней наступит утомление. Примером последействия может служить хорошо всем известная жизненная ситуация, когда даже после прекращения действия сильного эмоционального раздражителя (после прекращения ссоры) еще какое-то более или менее продолжительное время продолжается общее возбуждение, артериальное давление остается повышенным, сохраняется гиперемия лица, тремор кистей.
Таким образом, последействие объясняется наличием следовой деполяризации. Если следовая деполяризация длительна, то на ее фоне в течение нескольких миллисекунд может возникать потенциал действия (ритмическая активность нейрона), вследствие чего сохраняется ответная реакция. Но это дает сравнительно короткий эффект последействия. Более длительное последействие связано с наличием кольцевых связей между нейронами. В них возбуждение как бы само себя поддерживает, возвращаясь по коллатералям аксонов к первоначально возбужденному нейрону. От кольцевых связей между нейронами зависит электрическая активность нервных центров, имеющая место и при отсутствии афферентных стимулов, которую называют тонусом центров. Это явление проявляется в том, что нервный центр вследствие спонтанной активности посылает к органам и тканям редкие импульсы и обусловливает их тонус (тонус скелетных мышц, гладких мышц внутренних органов, стенок сосудов и др.). Тоническое состояние центров поддерживается различными гуморальными факторами и афферентными импульсами.
6. Нервные центры обладают высокой чувствительностью к недостатку кислорода. Нервные клетки отличаются интенсивным потреблением кислорода. Мозг человека поглощает около 40–70 мл кислорода в минуту, что составляет 1/4–1/8 часть всего количества кислорода, потребляемого организмом. Потребляя большое количество кислорода, нервные клетки высокочувствительны к его недостатку. Полное или частичное прекращение кровообращения центра (при тромбозе или разрыве кровеносных сосудов) ведет к тяжелым расстройствам деятельности нейронов, а полное прекращение – к гибели в течение 5–6 мин. Даже кратковременная остановка мозгового кровообращения или кратковременное резкое падение давления в кровеносных сосудах головного мозга вызывает у человека немедленную потерю сознания. Особенно сильно страдают при прекращении кровоснабжения клетки коры больших полушарий головного мозга. Центры ствола мозга восстанавливают свои функции через
15–20 мин, а центры спинного мозга – через 20–30 мин полного прекращения притока к ним крови.
7. Нервные центры, как и синапсы, обладают высокой чувствительностью к действию различных химических веществ, особенно ядов (стрихнин, морфин, фенамин, кортизол, хлороформ, барбитураты, алкоголь). На одном нейроне могут располагаться синапсы, обладающие различной чувствительностью к различным химическим веществам. Поэтому можно подобрать такие химические вещества, которые избирательно будут блокировать одни синапсы, оставляя другие в рабочем состоянии. Это делает возможным корректировать состояния и реакции как здорового, так и больного организма. Некоторые вещества действуют на определенные нервные центры. Например, апоморфин влияет болеет резко на рвотный центр, а лобелин – на дыхательный. Стрихнин блокирует функцию тормозных синапсов и вызывает резкое повышение возбудимости ЦНС, особенно спинного мозга. Кардиазол действует избирательно на двигательную зону больших полушарий.
8. Нервные центры, как и синапсы, обладают быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми. Это обусловлено резким уменьшением запасов медиатора в нервных окончаниях, уменьшением чувствительности к медиатору постсинаптической мембраны нервной клетки, уменьшением ее энергетических запасов, что наблюдается при длительной работе и является основной причиной развития утомления. Кроме того, в процессе деятельности нейрона наступает постепенное снижение чувствительности его рецепторов к медиатору, что называется десенситизацией. Быстрая утомляемость нервного центра проявляется в постепенном снижении и в конечном итоге полном прекращении рефлекторного ответа при продолжительном раздражении афферентных нервных волокон.
9. Нервные центры, как и синапсы, обладают низкой лабильностью, основной причиной которой является синаптическая задержка. Суммарная синаптическая задержка, наблюдающаяся во всех нейро-нейрональных синапсах при проведении импульсации по ЦНС, или в нервном центре, называется центральной задержкой.
10. Нервные центры обладают тонусом, который выражается в том, что даже при отсутствии специальных раздражений они постоянно посылают импульсы к рабочим органам. Некоторые рефлекторные акты протекают в течение длительного времени без видимого утомления. Это, например, тонические рефлексы с длительным, многочасовым поддержанием мышечного тонуса. При осуществлении того или иного рефлекса даже в состоянии относительного покоя из нервных центров на периферию к соответствующим органам и тканям поступают разряды нервных импульсов.
При осуществлении рефлекторной реакции они более частые и идут по большему числу эфферентных волокон.
Редкие импульсы, непрерывно поступающие из нервных центров на периферию, обусловливают тонус скелетных мышц, тонус мышц кишечника, сосудистый тонус.
Такое постоянное возбуждение нервных центров и называется тонусом нервных центров. Тонические рефлексы – обязательные спутники всякого двигательного акта. Особенно велико их значение в передвижении тела в пространстве, т.е. локомоции.
11. Нервные центры обладают пластичностью – способностью изменять собственное функциональное назначение и расширять свои функциональные возможности. Также пластичность можно определить как способность одних нейронов брать на себя функцию пораженных нейронов того же центра. Именно с явлением пластичности связана способность восстанавливать двигательную активность конечностей, например, ног, утраченную в результате травм спинного мозга. Однако это возможно только при поражении части нейронов данного центра или сохранении целостными части проводящих путей ЦНС. При полном разрыве спинного мозга восстановление двигательной активности оказывается невозможным. Кроме того, нейроны одного центра, например сгибателей, не могут брать на себя функцию нейронов другого центра – разгибателей, т.е. явление пластичности центров ЦНС ограничено.
Таким образом, посттравматическая пластичность нейронных бъединений выполняет компенсаторную (восстановительную) функцию, а пластичность, вызванная длительным афферентным раздражением – приспособительную функцию. Например, для процесса обучения пластичность нейронных объединений является необходимым условием, т.е. его рабочим механизмом.
12. Окклюзия (запирание, закупорка) – (лат. осclusus – запертый) осуществляется (также как и пространственная суммация) в конвергирующей системе соединения нейронов. Одновременной активации нескольких (минимум двух) рецепторов сильным или сверхсильным раздражителями к одному нейрону будут конвергировать несколько пороговых или сверхпороговых импульсов. На этом нейроне будет происходить окклюзия, т.е. эти два раздражителя ответят с той же максимальной силой, что и на каждый из них в отдельности. Феномен окклюзии состоит в том, что количество возбужденных нейронов при одновременном раздражении афферентных входов обоих нервных центров оказывается меньше, чем арифметическая сумма возбужденных нейронов (реакций) при раздельном (изолированном) раздражении каждого афферентного входа в отдельности (рецептивных полей).
Причина феномена окклюзии – перекрытие путей на вставочных или эфферентных нейронах с помощью конвергенции.
Явление окклюзии приводит к снижению силы ответной реакции. Окклюзия имеет охранительное значение, предотвращая перенапряжение нейронов при действии сверхсильных раздражителей.
13. Индукция в нервных клетках заключается в наведении противоположного процесса (торможения) возбуждением, и наоборот. Индукция бывает одновременной и последовательной. Одновременная – наблюдается в нескольких центрах одновременно: в одном, например, возбуждение, а в соседних – торможение. Последовательная индукция наблюдается в одном и том же центре, где вслед за возбуждением наводится торможение.
Различают положительную и отрицательную индукции. Положительная – когда наводится возбуждение вслед за торможением или в соседних центрах, или в одном центре при одновременном торможении одних центров. А под отрицательной
индукцией понимают такую, когда возбуждение наводится
торможением или в соседних центрах, или в том же, вслед за
возбуждением.
14. Облегчение проведения, или проторение пути. Установлено, что после возбуждения, возникшего в ответ на ритмическое раздражение, следующий стимул вызывает больший эффект или для поддержания прежнего уровня ответной реакции требуется меньшая сила последующего раздражения. Это явление получило название облегчения. Его можно объяснить тем, что при первых стимулах ритмического раздражителя происходит перемещение пузырьков медиатора ближе к пресинаптической мембране и при последующем раздражении медиатор быстрее выделяется в синаптическую щель. Это, в свою очередь, приводит к тому, что вследствие суммации ВПСП быстрее достигается критический уровень деполяризации и возникает распространяющийся потенциал действия.
Координация рефлекторной деятельности ЦНС.
Основные принципы
Координация рефлекторной деятельности ЦНС представляет собой согласованную работу нейронов ЦНС, основанную на взаимодействии нейронов между собой и одновременной реализации многих свойств ЦНС. Координация происходит от слова «сoordinati», что означает упорядочение. Основная функция нервной системы заключается в приспособлении организма к окружающим условиям. Но это возможно лишь тогда, когда рефлекторная деятельность упорядочена. Так, при раздражении любого участка тела должна быть диффузная реакция всех мышц тела и конечностей. Это происходит в силу анатомического строения ЦНС, обеспечивающего соединение различных нейронов не только на одном уровне, но и расположенных в ее верхних и нижних этажах (наблюдается у новорожденных младенцев). У взрослого человека рефлекторная деятельность упорядочена: если уколоть какой-либо палец правой руки, то отдергивается правая рука, а не левая, и тем более не нога.
Центральное торможение
 Явление центрального торможения было открыто И.М. Сеченовым в 1862 г. в эксперименте на таламической лягушке, у которой перерезали головной мозг на уровне зрительных бугров и удаляли полушария головного мозга (рис. 13). Затем по методу Тюрка измеряли время рефлекса отдергивания задних лапок (сгибательного рефлекса) при погружении их в раствор соляной кислоты. Время рефлекса – это время от начала раздражения до начала ответной реакции. Затем на область зрительных бугров накладывался кристалл поваренной соли, и время рефлекса увеличивалось, т.е. в таламической области мозга у лягушки существуют нервные центры, оказывающие тормозящее влияние на спинномозговые рефлексы.
Явление центрального торможения было открыто И.М. Сеченовым в 1862 г. в эксперименте на таламической лягушке, у которой перерезали головной мозг на уровне зрительных бугров и удаляли полушария головного мозга (рис. 13). Затем по методу Тюрка измеряли время рефлекса отдергивания задних лапок (сгибательного рефлекса) при погружении их в раствор соляной кислоты. Время рефлекса – это время от начала раздражения до начала ответной реакции. Затем на область зрительных бугров накладывался кристалл поваренной соли, и время рефлекса увеличивалось, т.е. в таламической области мозга у лягушки существуют нервные центры, оказывающие тормозящее влияние на спинномозговые рефлексы.
Прекращение воздействия соли на зрительные бугры приводило к восстановлению исходного времени рефлекторной реакции.
Этот рефлекс осуществляется спинальными центрами, и его время служит показателем возбудимости нервных центров.
Кристаллик соли, раздражая зрительные бугры, вызывает возбуждение, которое распространяется к спинальным центрам и тормозит их деятельность. И.М. Сеченов пришел к заключению, что торможение является следствием взаимодействия двух или более возбуждений на нейронах ЦНС. В этом случае одно возбуждение становится тормозимым, другое – тормозящим.
Таким образом, И.М. Сеченов доказал, что наряду с торможением в периферической нервной системе существует и центральное торможение, характерное для ЦНС.
Торможение в ЦНС выполняет две основные функции. Во‑первых, оно координирует функции, т.е. направляет возбуждение по определенным путям к определенным нервным центрам, при этом выключая те пути и нейроны, активность которых в данный момент не нужна для получения конкретного приспособительного результата. Ес






