Как известно, в заднюю центральную извилину, расположенную в теменной доле, в так называемой постцентральной области коры, проецируются рецепторы, заложенные в органы движения (мышцы и связки) и приносящие в кору кинестетические ощущения, то есть ощущения от положения и движений тела, его частей, работы мышц, а также кожные ощущения. Именно задняя центральная извилина, если использовать терминологию И.П. Павлова, должна считаться двигательным анализатором в узком смысле слова, а по определению Г.И. Полякова (1959), она является кожно-кинестетическим анализатором, ядерную зону которого, по его мнению, составляют поля 3, 1, 2 (рис. 2). Как уже было отмечено выше, А.Р. Лурия называет эту область мозговой коры "сензорным" отделом в составе двух совместно работающих зон двигательного анализатора. Исходя из различий клинической картины при поражении указанных полей, Г.И. Поляков выделяет одно, занимающее центральное, или первичное, положение в ядерной зоне, таковым является поле 3 (выделено нами-В.Д.). Обычно при повреждении этого центрального поля наблюдаются, как правило, резко выраженные симптомы нарушений в восприятии ощущений наиболее тонких дифференцировок соответствующих двигательных и кожных раздражителей.
Обнаружено (Г.И. Поляков, 1959; И.И. Филимонов, 1957 и др.), что первичное поле 3, как и центральные поля других ядерных зон (зрительной, слуховой), резко выделяется своеобразием своей цитоархитектонической картины, заключающемся в особенностях нейронного строения, обеспечивающего двусторонние корково-подкорковые связи, наиболее прямым и коротким путем соединяющие кору с рецепторами.
В центральных полях, указывает Г.И. Поляков, особенно многочисленны зерновидные клетки слоя IV, переключающие импульсы, приходящие по мощно развитым афферентным проекционным волокнам из подкорковых отделов анализаторов, на пирамидные нейроны слоев III и V коры. Г.И. Поляков (1962) также отмечает, что важнейшей чертой структурной и функциональной организации первичного поля 3 является наличие в нем четкой соматотопической проекции, при которой отдельные точки периферии (кожной поверхности, скелетной мускулатуры тела) проецируются в строго ограниченные соответствующие точки первичных полей, получивших в связи с этим название "проекционных полей".
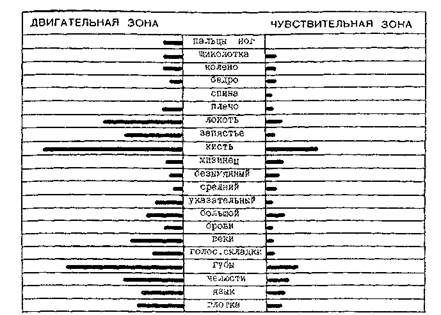
Установлена важная закономерность, что соматотопическая проекция строится в первичных полях коры не пропорционально величине частей и участков тела, а пропорционально их функциональному назначению. Так, наибольшей протяженностью и объемом отличаются участки коры, куда проецируются кожные и мышечные рецепции пальцев рук и кисти, органов артикуляции, характеризующиеся наиболее выраженной способностью дифференциации отдельных мельчайших раздражений (рис.1, рис.3).
В связи с указанной закономерностью Г.И. Поляков заключает, что "первичные поля ядерных зон представляют собой у человека высокоспециализированные участки коры, позволяющие весьма дробно выделять отдельные мельчайшие раздражители, что является, - по его мнению, - совершенно необходимым условием синтетического восприятия сложных комплексов раздражителей", приходящих от кожно-кинестетических рецепторов (А.Р. Лурия, 1962, С.43). Однако, замечает Г.И. Поляков, "наличие строго разграниченных отдельных пунктов проекций в пределах первичных полей следует понимать не как статическую, а как подвижную, динамическую конструкцию" (там же), что особенно отчетливо выступает в первичном двигательном поле коры (прецентральное гигантопирамидное поле 4). Это явление, как следует из литературы, доказано многочисленными опытами, в которых было обнаружено, что электрическое раздражение, приложенное к одной и той же точке двигательной коры, может давать различные эффекты в зависимости от разной интенсивности прикладываемого тока и предшествующего состояния данного коркового участка. На наш взгляд, эти данные имеют существенное значение в плане понимания патогенеза двигательных нарушений коркового происхождения в раннем онтогенезе, а также для формирования представлений о тех компенсаторных механизмах, которые могут обеспечить в этих случаях коррекцию нарушенных двигательных функций.
Установлено, что периферические, или вторичные, поля, расположенные в периферических отделах ядерных зон (для кожно-кинестетического анализатора это поля 1 и 2), отличаются от центральных, или первичных, полей как по особенностям архитектоники и нейронной организации, так и по функциональным проявлениям. Замечено, что при поражении или электрическом раздражении этих нолей возникают эффекты, касающиеся преимущественно более сложных форм психической (применительно к нашему контексту - психомоторной) деятельности.
При относительной сохранности элементарных ощущений (в нашем случае кинестетических) расстраивается главным образом способность воспринимать целые комплексы и взаимные соотношения элементов сложных раздражителей. В преломлении к психомоторике, как нам это представляется, в подобных случаях будет наблюдаться нарушение кинестетической основы сложных двигательных актов, что обусловит затруднения в формировании произвольной моторики, особенно гонких дифференцированных движений.
Рис. 3. Объем проекционных областей в коре головного мозга (по Л.О. Бадаляну, 1987)
Нейронная структура вторичных полей, как свидетельствуют нейрофизиологические исследования, приспособлена для переключения приходящих из подкорки афферентных импульсов через зерновидные клетки слоя IV на крупные пирамиды нижнего уровня слоя Ш, дающие начало наиболее мощной системе ассоциационных связей коры (вторичный проекционно-ассоциационный нейронный комплекс; рис.4, ). Эта особенность цитоархитектоники, по мнению Г.И. Полякова, обусловливает весьма существенную роль вторичных полей ядерных зон "в осуществлении взаимных связей отдельных раздражителей, выделяемых через посредство первичных полей, а также в функциональном объединении ядерных зон разных анализаторов в процессе интеграции сложных комплексов реакций, идущих по разным анализаторам" (А.Р. Лурия, 1962; С.44).
Из этого замечания следует, что вторичные поля имеют преимущественное отношение к осуществлению более сложных форм координированной психической деятельности, связанной с детальным анализом соотношений конкретных раздражителей и с ориентировкой в конкретном окружающем пространстве и времени. Именно вторичные
поля, как мы понимаем это замечание применительно к психомоторике, определяют тонкую координированную работу мышц и мышечных групп благодаря анализу поступающей в эти поля кинестетической информации or органов, задействованных в той или иной совместной психомоторной деятельности.
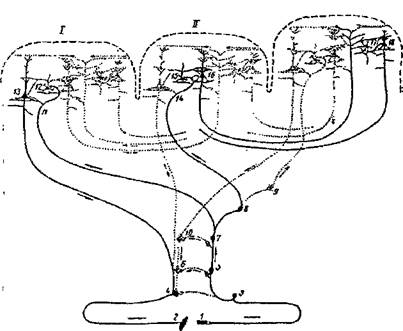
Рис.4. Система связей первичных, вторичных и третичных полей коры (по Г.И. Полякову, 1959)
/ - первичные (центральные) поля; // - вторичные (периферические) поля 111 - третичные поля (зоны перекрытия анализаторов).
Жирными линиями выделены
/ - система проекционных (корково-подкорковых) связей коры;
// - система проекционно-ассоциационных связей коры;
/// - система ассоциационных связей коры.
/— рецептор,
2 — эффектор.
3 —нейрон чувствительного узла;
4 — двигательный нейрон,
5 — б — переключательные нейроны спинного мозга и ствола;
7— 10 — переключательные нейроны подкорковых образований;
//, 14 — афферентное волокно из подкорки;
13 — пирамида V слоя,
16 — пирамида подслоя ПГ",
18 — пирамиды подслоев III" и ИГ;
12, 15, 17 — звездчатые клетки коры
Экспериментально методом нейронографии доказано, что возбуждение, возникшее в результате раздражения первичных полей коры, имеет лишь сравнительно ограниченное распространение непосредственно в пределах соответствующих ядерных зон, тогда как возбуждение, возникающее во вторичных полях, имеет тенденцию распространяться на значительно большие пространства коры за пределами определенной ядерной зоны. Тем самым проекционные связи вторичных полей коры характеризуются по сравнению с первичными большей сложностью всей системы переключений в подкорковых отделах анализаторов, что наглядно может быть представлено схемой Г.И. Полякова (рис.5). Как видно из схемы, афферентные импульсы от рецепторов к первичным и вторичным полям приходят разными путями: во вторичные поля через большее число дополнительных переключений в ассоциационных ядрах зрительного бугра, а в первичные поля более коротким путем через реле-ядра зрительного бугра.
Таким образом, по мнению Г.И. Полякова, "рецепции, достигающие периферических полей и используемые для широких ассоциаций как внутри соответствующих зон, так и между разными зонами коры уже в подкорковых уровнях анализаторов, проходят определенную степень предварительной интеграции, которая распространяется не только на систему данного анализатора, но и на разные анализаторы, кооперирующиеся между собой в процессе координированной совместной деятельности" (А.Р. Лурия, 1962; С.45). Исходя из приведенного замечания, становится вполне понятно, почему поражение или недоразвитие первичных или вторичных корковых полей анализаторов будет вызывать нарушения разного порядка: от специфичных симптомов, связанных с расстройством какого-либо определенного анализатора (в нашем случае двигательного), до сложных симптомокомплексов, указывающих на недостатки в функционировании нескольких анализаторов разной модальности.
В этой связи представления о различных путях прохождения аф-ферентационных импульсов от рецепторов к разным корковым полям и знание функциональной роли первичных и вторичных полей могут помочь, как нам представляется, разобраться в особенностях симптоматики того или иного двигательного нарушения, в структуре сложного дефекта и его механизмах, а также установить возможную локализацию мозгового поражения.
Как доказано многими работами отечественных и зарубежных исследователей, в процессе эволюционного расширения ядерной зоны кожно-кинестетического анализатора, равно как и аналогичных областей других анализаторов задних отделов мозга (зрительного и слухового), по поверхности полушария произошло взаимное их проникновение с образованием особых "зон перекрытия" корковых концов анализаторов, так называемых третичных полей.

Рис.5. Схема переключений в системах анализаторов (по Г.И.Полякову, 1959)
/ - периферические воспринимающие поверхности анализаторов;
2, 3, 4 - переключения в подкорковых отделах анализаторов.
7, Д /Д- как на Рис. 4.
Также установлено, что группы нейронов, из которых состоят третичные поля коры, по сравнению с комплексами нейронов ядерных зон (первичных и вторичных полей), почти полностью "освобождены" от специфических анализаторных функций. Доказано, что они (третичные поля) целиком переключены на адекватное отражение наиболее сложных форм пространственных и временных отношений между реальными комплексами раздражителей и на оперирование этими отношениями в процессе активного взаимодействия организма с внешним миром.
В этой связи повреждение или недоразвитие третичных полей не будет сопровождаться выраженными нарушениями специфических функций анализаторов, "...при поражении этих территорий расстраиваются главным образом наиболее обобщенные проявления корковой деятельности, опирающиеся на совместную работу нескольких анализаторов, позволяющие осуществлять наиболее сложные формы ориентировки во внешнем мире и анализ и синтез сложных систем отношений между раздражителями, воздействующими на воспринимающую часть разных анализаторов" (А.Р. Лурия, 1962; С.45).
Особенности микроструктуры и функциональное значение третичных полей определяются топографическими соотношениями этих полей с полями ядерных зон, между которыми они располагаются. Г.И.Поляков выделяет три частные зоны перекрытия в задних отделах коры: верхнюю теменную область (поле 39), нижнюю теменную область (поле 40) и височно-теменно-затылочную подобласть (поля 21 и 37), - расположенные на стыках вторичных полей ядерных зон анализаторов (рис.2 и рис.6). Как видно на схеме (рис.6), обе теменные области (верхняя - поле 39 и нижняя - поле 40), исходя из их расположения, топографически и функционально тесно связаны с кожно-кинестетическим и зрительным анализаторами.
Верхняя теменная область граничит спереди с теми отделами кожно-кинестетического анализатора, в которых представлены проекции hoi и туловища. Тем самым эта область, как доказано, имеет специальное значение для интеграции со зрительными рецепциями движений всего тела и в формировании схемы тела.
Нижняя теменная область граничит с теми отделами кожно-кинестетического анализатора, в котором представлены проекции рук и лица. Соответственно эта область, указывает Г.И. Поляков, "имеет отношение к интеграции обобщенных, и отвлеченных форм сигнализации, которые связаны с тонко и сложно дифференцированными предметными и речевыми действиями, совершаемыми под контролем зрения и требующими совершенно разработанной системы ориентировки в окружающем пространстве" (А.Р. Лурия, 1962; С.47; выделено нами- В.Д.).
Височно-теменно-затылочная подобласть, являясь зоной взаимоперехода слуховой и зрительной зон коры, как свидетельствуют клинические данные, функционально тесно связана с наиболее сложными формами интеграции слуховых и зрительных рецепций, а также с определенной стороной семантики устной и письменной речи (А.Р. Лурия, Г.И. Поляков и др.).
Рис.6. Схема корково-подкорковых соотношений первичных, вторичных и третичных зон (по Г.И.Полякову, 1959)
Жирными линиями показаны системы анализаторов с их переключениями в подкорковых отделах:
/ - зрительный анализатор;
2-слуховой анализатор;
3 -кожно-кинестетический анализатор.
Т- височная область;
О- затылочная область;
Pip - поле 39;
Pia - поле 40;
Pstc - постцентральная область;
ТРО -височно-теменно-затылочная подобласть;
77? - зрительный бугор;
Cgm- внутреннее коленчатое тело;
Cgl - наружное коленчатое тело. (Обозначение полей 9, 10, 46 третичной зоны префронтальной области нанесено нами - В.Д.)
Установлено, что в архитектонической и нейронной организации зон перекрытия на первый план выступает третичный ассоциационныи комплекс коры (рис. 4, III). Он представлен, как выяснено, пирамид-

ными и звездчатыми клетками наиболее высоких по уровню расположения среднего и верхнего подслоев слоя III и слоя II.
Вместе с дифференцированными многосторонними ассоциационными связями третичные поля характеризуются по сравнению с первичными и вторичными полями и наиболее сложными цепями проекционных переключений в подкорковых отделах анализаторов.
В соответствии с наиболее высоким уровнем сообщения и отвлечения от конкретных реальных раздражителей третичные поля связаны с периферическими концами анализаторов наиболее опосредованным образом через наибольшее число последовательных переключений в подкорковых образованиях (рис. 4). В этой связи Г.И. Поляков делает вывод о том, что импульсы, подаваемые снизу к третичным полям задних отделов коры, "проходят наиболее сложную переработку в подкорке и подводятся к коре, как интегрированные сигналы, образовавшиеся в результате взаимодействия чувственных показаний различных анализаторов" (А.Р. Лурия, 1962; С.47).
В нормально действующем мозге все "три группы корковых полей вместе с их системами переключений и связей между собой и с подкорковыми инстанциями работают как одно сложнодифференцированное целое" (там же). Согласованная работа анализаторных систем задних отделов мозга (кожно-кинестетической, зрительной и слуховой), как надо понимать, и обеспечивает сенсорную в широком смысле слова основу психомоторной деятельности человека.
Следующий раздел посвящен аналогичному рассмотрению структурно-функциональных особенностей моторного отдела двигательного анализатора человека.






