ЦИТОЛОГИЯ. КЛЕТКА И НЕКЛЕТОЧНЫЕ СТРУКТУРЫ
Клетка -Основой строения эукариотических организмов является клетка (cellula) - наименьшая единица живого.
Эукариотические, собственно ядерные организмы — основная масса животных и растений, за исключением бактерий и сине-зеленых водорослей, не имеющих оформленного ядра — прокариотических организмов.
Клетка — элементарная структурная, функциональная и генетическая единица в составе всех растительных и животных организмов, участвующих в единой совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Каждая клетка включает в себя три основных компонента;
1. Плазмолемму
2. Цитоплазму
3. Ядро
ПЛАЗМОЛЕММА - Барьерно-рецепторная и транспортная система клетки.
Плазмолемма (plasmalemma ), или внешняя клеточная мембрана, среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой, а следовательно, и со всеми веществами и стимулами, воздействующими на клетку.
Плазмолемма обладает следующими свойствами:
· избирательной проницаемостью через пассивный и активный транспорт и диффузию;
· текучестью или подвижностью, многие белки могут скапливаться в одном участке мемраны, образуя агрегаты, тем самым осуществляется более эффективная рецепция.
Функции плазмолеммы. Эта мембрана выполняет ряд важнейших клеточных функций, ведущими из которых являются барьерная функция (разграничения цитоплазмы с внешней средой):
· Транспорт веществ в цитоплазму и из нее;
· Рецепция;
· Образование межклеточных контактов;
· Передача сигналов от поверхности в глуб клетки (с помощью сигнальных молекул – гормоны, медиаторы, цитокины).
· Движение клетки
СОСТАВ ПЛАЗМОЛЕММЫ:
Надмембранный комплекс
Мембранный комплекс
Субмембранный комплекс
1.1. Надмембранный комплекс – представлен гликокаликсом (glycocalyx), который имеет толщину около 50 нм. Гликокалис состоит из Олигосахаридов, связанных с белками (гликопротеины) и Олигосахаридов, связанных с липидами (гликолипиды).
Функции Гликокаликса:
1. Рецепсия и адгезия (слипание) клеток:
· рецепторы гистосовместимости (HLA –рецепторы);
· специфические рецепторы к гормонам;
· специфические рецепторы к медиаторам
· специфические рецепторы к цитокинам;
2. Межклеточные взаимодействия:
· воспринимает физические раздражения (например, кванты светав фоторецепторах);
· химические раздражения (вкусовые и обонятельные изменения рН);
· механические раздражения (давление, растяжение);
· узнавание.
3. Пристеночное пищеварение:
· гликокалис, покрывающий микроворсинки каемчатых клеток эпителия кишечника.
1.2. Мембранный комплекс – представлен элементарной биологической мембраной, котрая состоит из бислоя Фосфолипидов и встроенных в него белков.
Фосфолипиды состоят из Полярной (гидрофильной) головки и Неполярного (гидрофильного) хвоста. В мембране хвосты направлены в глубь бислоя, а головки обращены наружу.
Классификация липидов мембран:
· фосфолипиды - из мемранных фосфолипидов освобождается арахидоновая кислота, которая является предшественником простогландинов, тромбоксанов, лейкотриенов и других БАВ;
· сфинголипиды – чаще встречаются в миелиновой оболочке нервных волокон и швановских клетках;
· холестерин (холестерол) – играет важную роль, т.к. он является родоначальником синтеза стероидных гормонов – половых, глюкокортикойдов, минералокортикойдов, а также входит в состав липопротеидов (липопротеины низкой плотности ЛНП, липопротеины высокой плотности – ЛВП, изменение соотношения между которыми может повысить риск развития ишемической болезни сердца).
Классификация структурных белков, входящих в мембрану:
1. Интегральные – пронизывают всю мембрану насквозь. Функции: белки ионных каналов и мебранные рецепторы.
2. Полуинтегральные – (поверхностные):
· периферические белки наружной поверхности. Функция: рецепторная, адгезивная;
· периферические белки внутренней поверхности. Функция: участвуют в построении цитоскелета клеток (белки цитоскелета – спектрин, анкирин).
Классификация белков по выполняемым функциям:
· Белки - рецепторы.
· Белки – ферменты.
· Белки – переносчики
· Структурные белки
· Трансмембранные белки – интегрины – клеточные адгезионные молекулы (КАМ) – они являются рецепторами для фибронектина и ламина.
1.3. Субмембранный комплекс – представлен элементами опорносократительного аппарата клетки – ЦИТОСКЕЛЕТА
Цитоскелет (cytoskeleton) -опорно-двигательная система клетки, включающая немембранные белковые нитчатые органеллы, выполняющие как каркасную, так и двигательную функции в клетке. Эти структуры являются динамическими образованиями, они могут быстро возникать в результате полимеризации их элементарных молекул и так же быстро разбираться, исчезать при деполимеризации. К этой системе относятся фибриллярные структуры и микротрубочки.
Фибриллярные структуры цитоплазмы. К фибриллярным компонентам цитоплазмы эукариотических клеток относятся:
· Микрофиламенты (micro-filamenti) толщиной 5-7 нм. Микрофиламенты встречаются практически во всех типах клеток. Они располагаются в кортикальном слое цитоплазмы, непосредственно под плазмолеммой, пучками или слоями. Их можно видеть в псевдоподиях амеб или в движущихся отростках фибробластов, в микроворсинках кишечного эпителия. Микрофиламенты часто образуют пучки, направляющиеся в клеточные отростки.
· Промежуточные филаменты (filamenti intermedii) (рис. 4.16).Это тонкие (10 нм) неветвящиеся, часто располагающиеся пучками нити. Характерно, что в клетках разных тканей их белковый состав различен. Например, в эпителии кожного типа в состав промежуточных филаментов входит кератин. Пучки кератиновых промежуточных филаментов в эпителиальных клетках образуют тонофиламенты, которые подходят к десмосомам. В состав промежуточных филаментов клеток, производных мезенхимы (например, фибробластов), входит другой белок - виментин; в мышечных клетках обнаруживается десмин; в нервных клетках в состав нейрофиламентов также входит особый белок. Роль промежуточных микрофиламентов, скорее всего, опорно-каркасная; эти фибриллярные структуры не так лабильны, как микротрубочки и микро-филаменты.
В клинике с помощью иммуноморфологических методов тканевое происхождение тех или иных опухолей определяется именно по белкам их промежуточных филаментов. Это очень важно для диагностики и правильного выбора типа химиотерапевтических противоопухолевых препаратов.
· Микротрубочки –полые цилиндры, стенка которых стоит молекул белка – Тубулина;
· Миофиламенты – состоят из белка Миозина, выполняют сократительную функцию.
МЕМРАННЫЙ ТРАНСПОРТ
1. Пассивный транспорт – движение молекул в обоих направлениях по градиенту концентрации без затрат энергии. Различают:
· Простую;
· Облегченную диффузию.
Простая диффузия – характеризуется низкой специфичностью. Она осуществляет транспорт О2, СО2, N2, H2O. (Например, внешнее дыхание через аэрогематический барьер, или тканевое дыхание).
Облегченная диффузия – осуществляется с участием компонентов мембраны через ионные каналы или белки – переносчики. Проявляет специфичность по отношению к транспортируемым молекулам.
Ионные каналы – состоят из связанных между собой трансмембранных белков, формирующих в мембране небольшую пору. Через пору по электрохимическому градиенту проходят ионы. Наиболее распространенные каналы для Na+, K+, Ca+, Cl -.
Натриевые каналы -присутствуют в возбудимых структурах (например, скелетные мышечные волокна, кардиомиоциты, нейроны). Генерируют потенциал действия, осуществляют начальный этап деполяризации мембраны.
Калиевые каналы – находятся в плазмолемме всех клеток.Участвуют в поддержании мембранного потенциала, регулируютобьем клетки, моделируют электронную возбудимость нервных мышечных структур.
Кальцевые каналы:
· участвуют в депонировании Ca2+ в цистернах гладкой ЭПС;
· принимает участие в мышечном сокращении, секреции гормонов и нейромедиаторов и множестве других клеточных процессов.
· Хлорные каналы – присутствуют в плазмолемме скелетных мышечных волокон, регулируют электронную возбудимость плазмолемму. Уменьшение Сl+ - проводимости ведет к электронной нестойкости мембраны мышечных волокон и развитию миотонии.
· Водные каналы (аквапорины)- семейство мембранных пор для воды.
Ионные каналы состоят из собственно транспортной системы и воротного механизма, который открывает канал на некоторое время в ответ на изменение мембранного потенциала, механическое воздействие (волосковые клетки внутреннего уха, тельца Фатер-Пачинни), действия нейромедиаторов, биогенных аминов, АТФ, циклических нуклеотидов.
2. АКТИВНЫЙ ТРАНСПОРТ
- энергозависимый трансмембранный перенос против градиента концентрации с участием Na+, H+, Ca2+, ATФаз.
Примером механизма активного транспорта служит натриево-калиевый насос, осуществляемый с помощью белка – переносчика Na+, K+ - ATФазы (выкачивает Na+ из клетки в обмен на K+), который поддерживает постоянство обмена клеток и мембранный потенциал.
Другой пример – протонная и калиевая АТФаза, при помощи которой париентальные клетки желез слизистой оболочки желудка участвуют в образовании соляной кислоты.
3.ЭНДОЦИТОЗ – транспорт макромолекул в клетку.
варианты эндоцитоза:
· пиноцитоз
· фагоцитоз
· опосредственный рецепторами эндоцитоз
ПИНОЦИТОЗ – процесс поглощения жидкости и растворенных веществ с образованием эндоцитозных пузырьков (эндосом). Диаметр эндосом – 0.2- 0.3 мкм – макропиноцитоз; диаметр эндосом – 70-100 нм –микропиноцитоз.
ФАГОЦИТОЗ – поглощение крупных частиц (более 1 мкм) (микроорганизмов, или остатков клеток). Фагоцитоз осуществляется специальными клетками –фагоцитами (макрофаги, нейтрофиллы). В ходе фагоцитоза образуются фагосомы, при слиянии их с лизосомами образуются фаголизосомы.
Опосредованный рецепторами эндоцитоз -осуществляется через образование окаймленных пузырьков. Мембранные рецепторы макромолекул накапливаются в области эндоцитозных ямок. Вокруг таких ямок со стороны цитоплазмы образуется оболочка, состоящая из белка клатрина (имеет вид щетинистой каемки). Скопление рецепторов в одном месте создает условия к более эффективному эндоцитозу поглощаемого вещества (лиганда). Переваривание (процессинг) внутри окаймленного пузырька происходит только только после того, как разрушается клатриновая оболочка. Если она не утрачивается, то содержимое пузырька остается не изменным. Окаймленные пузырьки транспортируют иммуноглобулины, белки желточных включений, факторы роста, трансферрин, липопротеины низкой плотности (ЛНП).
4.ЭКЗОЦИТОЗ – процесс при котором внутриклеточные секреторные гранулы сливаются с плазмолеммой, и их содержимое освобождается из клетки – секреция во внеклеточное пространство.
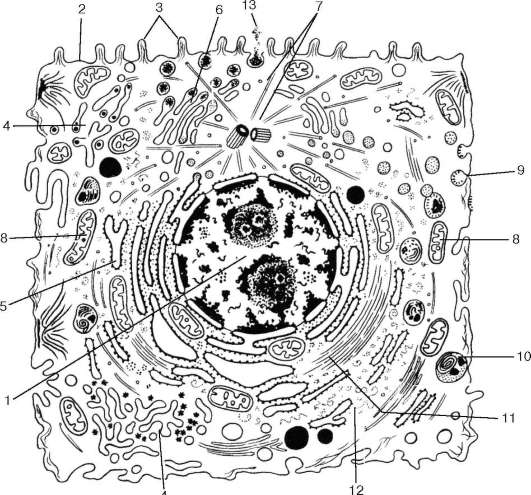
Рис. 4.1. Ультрамикроскопическое строение клетки животных организмов (схема): 1 - ядро; 2 - плазмолемма; 3 - микроворсинки; 4 - агранулярная эндоплазматическая сеть; 5 - гранулярная эндоплазматическая сеть; 6 - комплекс Гольджи; 7 - центриоль и микротрубочки клеточного центра; 8 - митохондрии; 9 - цито-плазматические пузырьки; 10 - лизосомы; 11 - микрофиламенты; 12 - рибосомы; 13 - выделение гранул секрета
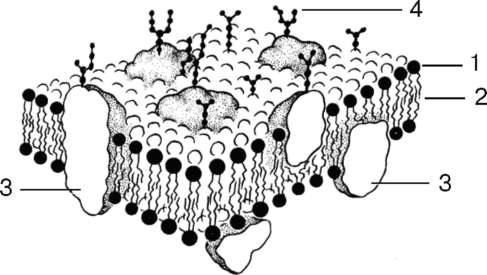
Рис. 4.2. Строение клеточной мембраны (схема);
СТРУКТУРНЫЕ КОМПОНЕНТЫ
ЦИТОПЛАЗМА
Цитоплазма (cytoplasma), Цитоплазма, часть клетки, отделенная от окружающей среды плазмолеммой, включает - Гиалоплазму (Содержимое цитоплазмы, лишенное органелл и включений, называют Гиоплазмой) и находящиеся в ней обязательные клеточные компоненты - Органеллы, а также различные непостоянные структуры - Включения.
1. ГИОПЛАЗМА ( цитозоль, матрикс) – внутренняя среда клетки, составляющая 55% от общего обьема клеткиПредставляет собой коллоидную систему, которая претерпевает превращения по типу гель-золь. Включает в себя биополимеры: белки, нуклеиновые кислоты, полисахариды, ферменты метаболизма сахаров, аминокислот, липидов.
Гиоплазма обьединяет все органеллы клетки, здесь синтезируются цитозольные белки, происходит переваривание короткоживущих белков с помошью нелизосомальных протеаз, осуществляются транспортные прпоцессы (элементы цитоскелета), запасающая функция (включения).
2. Органеллы
Органеллы - постоянно присутствующие и обязательные для всех клеток микроструктуры, выполняющие жизненно важные функции.
Классификация органелл. Различают:
* мембранные и
* немембранные органеллы.
Мембранные органеллы представлены:
- цитоплазматической сетью (эндо-плазматическим ретикулумом),
- комплексом Гольджи (аппаратом Гольджи),
- митохондриями,
- лизосомами,
- пероксисомами.
К немембранным органеллам относят:
- рибосомы (полирибосомы),
- клеточный центр и
- элементы цитоскелета (микротрубочки, микрофиламенты и промежуточные филаменты).
МЕМБРАННЫЕ ОРГАНЕЛЛЫ:
· Гранулярная эндоплазматическая сеть (reticulum endoplasmicum granulosum) (открыта К.Портер,1945), на ультратонких срезах представлена замкнутыми мембранами, которые на сечениях образуют уплощенные мешки, цистерны, трубочки.
Диаметр цистерн значительно варьирует и в зависимости от функциональной активности клетки колеблется от 20 нм до несколько микрометров. Отличительной чертой мембран гранулярной эндоплазматической сети является то, что они со стороны гиалоплазмы покрыты многочисленными рибосомами (рис.4.10). Гранулярная эндоплазматическая сеть имеет разное строение. Для малоспециализированных клеток или для клеток с низкой метаболической активностью характерно наличие редких и разрозненных цистерн. Если возникают локальные скопления гранулярной эндоплазматической сети, то это свидетельствует об активном синтезе секреторных белков. Так, в клетках печени и некоторых нервных клетках гранулярная эндоплазматическая сеть собрана в отдельные зоны.
В клетках поджелудочной железы гранулярная эндоплазматическая сеть в виде плотно упакованных друг около друга мембранных цистерн занимает базальную и околоядерную зоны клетки. Рибосомы, связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков - ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях эндоплазматической сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они модифицируются и входят в состав либо лизосом, либо секреторных гранул, содержимое которых остается изолированным от гиалоплазмы мембраной. Внутри канальцев или вакуолей гранулярной эндоплазматической сети происходит модификация белков, например, связывание их с сахарами (первичное глюкозилирование).
В гранулярной эндоплазматической сети на ее рибосомах происходит синтез мембранных интегральных белков, которые встраиваются в толщу мембраны. Здесь же со стороны гиалоплазмы идет синтез липидов и их встраивание в мембрану. В результате этих двух процессов наращиваются сами мембраны эндоплазматической сети и другие компоненты вакуоляр-ной системы клетки.
Итак, роль гранулярной эндоплазматической сети заключается в синтезе на ее рибосомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, в химической модификации таких белков и в их локальной конденсации, а также в синтезе структурных компонентов клеточных мембран.
· Комплекс Гольджи
Комплекс Гольджи (пластинчатый комплекс) (был открыт в 1898 г. К. Гольджи). Автор, используя свойства связывания тяжелых металлов (осмия или серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал внутренним сетчатым аппаратом (apparatus reticularis internus). В дальнейшем его стали называть аппаратом, или комплексом Гольджи (complexus Golgiensis). Подобные структуры затем были описаны во всех клетках эукариот. При рассмотрении в электронном микроскопе комплекс Гольджи представлен мембранными структурами, собранными вместе в небольших зонах (рис. 4.11).
Отдельная зона скопления этих мембран называется диктиосомой (стопкой Гольджи). Таких зон в клетке может быть несколько. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены 5-10 плоских цистерн, между которыми находятся тонкие прослойки гиалоплазмы. Каждая цистерна имеет переменную толщину: в центре ее мембраны могут быть сближены (до 25 нм), а на периферии иметь расширения - ампулы, ширина которых непостоянна. Кроме плотно расположенных плоских цистерн, в зоне комплекса Гольджи наблюдается множество мелких пузырьков (везикул), которые встречаются главным образом в его периферических участках. Иногда они отшнуровываются от ампулярных расширений на краях плоских цистерн. В зоне диктиосомы различают проксимальную (cis) и дистальную (trans) поверхности. В секретирующих клетках обычно комплекс Гольджи поляризован: его проксимальная поверхность обращена к ядру, в то время как дистальная - к поверхности клетки.
Комплекс Гольджи участвует в сегрегации и накоплении продуктов, синтезированных в эндоплазматической сети, в их химической перестройке, созревании; в его цистернах происходят синтез полисахаридов, их ком-плексирование с белками, что приводит к образованию пептидоглика-нов. С помощью комплекса Гольджи осуществляется процесс выведения готовых секретов за пределы секреторной клетки. Кроме того, комплекс Гольджи обеспечивает формирование лизосом. Мембраны комплекса образуются путем отщепления мелких вакуолей от гранулярного эндоплазматического ретикулума. Эти вакуоли поступают в проксимальный отдел комплекса Гольджи, где и сливаются с его мембранами.
Следовательно, в комплекс Гольджи поступают новые порции мембран и продуктов, синтезированных в гранулярном эндоплазма-тическом ретикулуме. В мембранных цистернах комплекса Гольджи происходят вторичные изменения в структуре белков, синтезированных в гранулярном эндоплазматическом ретикулуме. Эти изменения (модификации) связаны с перестройкой олигосахаридных цепочек синтезированных гликопротеидов. Внутри полостей комплекса Гольджи с помощью различных ферментов (транс-глюкозидаз) по разному модифицируются лизосомные белки и белки секретов: происходят последовательная замена и наращивание олигосахаридных цепочек. Модифицирующиеся белки переходят от цистерны проксимальной цис-поверхности в цистерны дистальной поверхности путем эстафетного переноса мелких вакуолей, содержащих транспортируемый белок.
· Митохондрии (mitochondrial) -энергетическая система клетки, органеллы синтеза АТФ. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ. Исходя из этого, митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания.
Термин «митохондрия» был введен Бенда в 1897 г. для обозначения зернистых и нитчатых структур в цитоплазме разных клеток. Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. Форма и размеры митохондрий животных клеток разнообразны, но в среднем толщина их около 0,5 мкм, а длина - от 1 до 10 мкм. Подсчеты показывают, что количество их в клетках сильно варьирует - от единичных элементов до сотен. Так, в клетке печени они составляют более 20 % общего объема цитоплазмы и содержат около 30-35 % общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазматической мембраны.
· 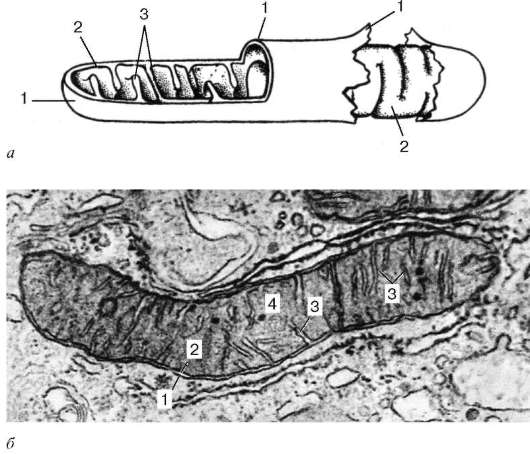
Рис. 4.14. Ультрамикроскопическое строение митохондрии:
А - схема; б - электронная микрофотография среза митохондрии печеночной клетки. 1 - наружная митохондриальная мембрана; 2 - внутренняя митохондриальная мембрана; 3 - кристы; 4 - митохондриальный матрикс
· Лизосомы (lysosomae) – (были открыты в 1949 г. де Дювом), -это разнообразный класс вакуолей размером 0,2- 0,4 мкм, ограниченных одиночной мембраной. Характерным признаком лизосом является наличие в них гидролитических ферментов - гидролаз (протеиназы, нуклеазы, фосфатазы, липазы и др.), расщепляющих различные биополимеры при кислом значении рН.
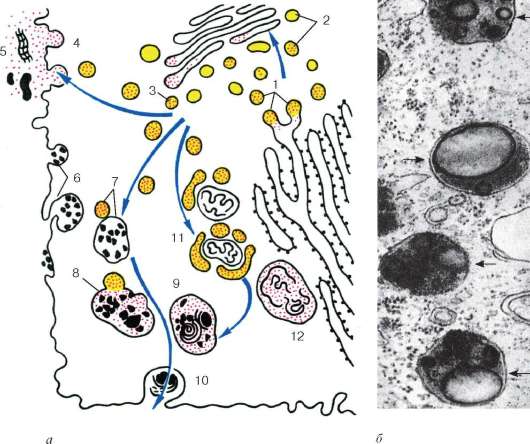
Рис. 4.13. Строение лизосом:
а - схема участия структур клетки в образовании лизосом и во внутриклеточном пищеварении: 1 - образование из гранулярной эндоплазматической сети мелких пузырьков, содержащих гидролитические ферменты; 2 - перенос ферментов в комплекс Гольджи; 3 - образование первичных лизосом; 4 - выделение и использование (5) гидролаз при внеклеточном расщеплении; 6 - эндоцитозные пузырьки; 7 - слияние первичных лизосом и эндоцитозных пузырьков; 8 - образование вторичных лизосом; 9 - телолизосомы; 10 - экскреция остаточных телец; 11 - слияние первичных лизосом с разрушающимися структурами клетки; 12 - аутофаголизосома; б - электронная микрофотография среза гетерофаголизо-сом (обозначены стрелками)
К немембранным органеллам относят:
· Рибосомы (ribosomae) - элементарные аппараты синтеза белковых, полипептидных молекул - обнаруживаются во всех клетках (рис. 4.15). Рибосомы - это сложные рибону-клеопротеиды, в состав которых входят белки и молекулы рибосо-мальных РНК (рРНК) примерно в равных весовых отношениях. Размер функционирующей рибосомы эукариотических клеток 25x20x20 нм. Такая рибосома состоит из большой и малой субъединиц. Каждая из субъединиц построена из рибонуклеопро-теидного тяжа, где рРНК взаимодействует с разными белками и образует тело рибосомы.
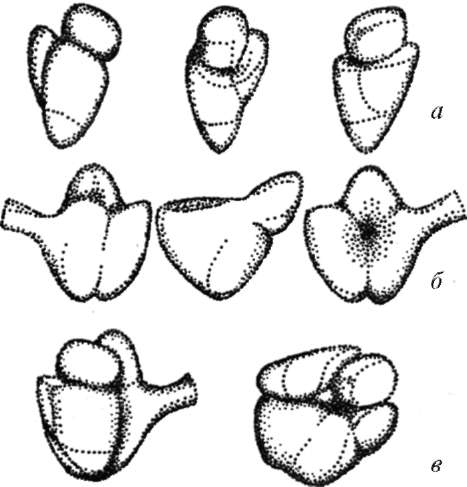
Рис. 4.15 Строение рибосом:
А - схема; б - электронная микрофотография. 1 - активная материнская центриоль, окруженная тонкофибриллярным матриксом, от которого отходят микротрубочки полярной лучистости (2); 3 - неактивная дочерняя центриоль
Межклеточные соединения
Плазмолемма принимает активное участие в образовании специальных структур - межклеточных контактов, или соединений (junc-tiones intercellulares), обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур (рис. 4.4).
Общим для этих клеток является то, что на их поверхности располагаются специальные углеводные части интегральных белков, гликопротеидов, которые специфически взаимодействуют и соединяются с соответствующими белками на поверхности соседних клеток.
Рис. 4.9. Щелевое соединение: (1 - коннексон; 2 – плазмолемма
Коммуникационное)
.
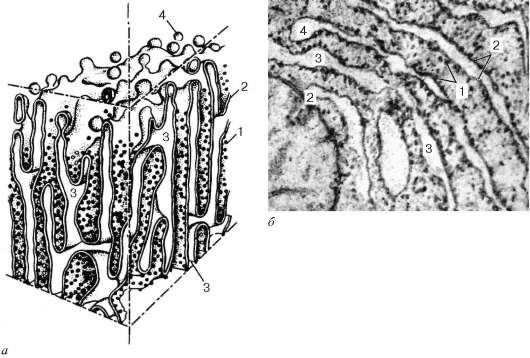
Рис. 4.10. Строение гранулярной эндоплазматической сети: а - схема; б - электронная микрофотография участка среза эпителиальной клетки печени. 1 - рибосомы; 2 - пластинки; 3 - внутренние полости цистерн; 4 - отщепляющиеся мембранные пузырьки, лишенные рибосом
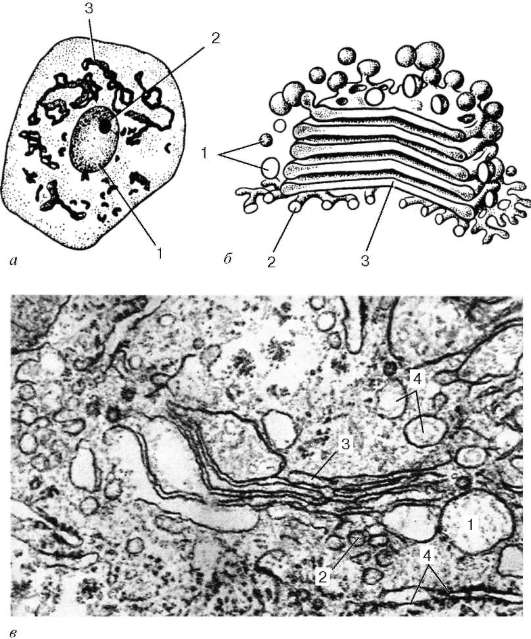
Рис. 4.11. Комплекс Гольджи:
а - нервная клетка спинного мозга, импрегнация серебром по методу Гольджи: 1 - ядро; 2 - ядрышко; 3 - комплекс Гольджи; б - схема ультрамикроскопического строения (трехмерная реконструкция); в - комплекс Гольджи на ультратонком срезе (печеночная клетка): 1 - пузырьки; 2 - трубочки; 3 - уплощенные мешочки (цистерны); 4 - фрагменты гранулярной эндоплазматической сети
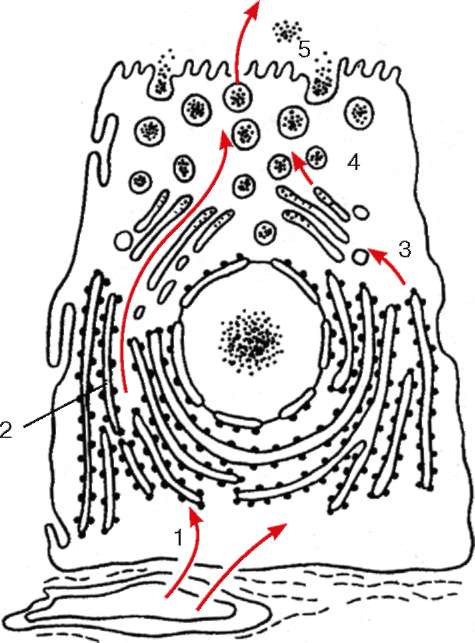
Рис. 4.12. Участие клеточных структур в белковой секреции (схема): 1 - поступление аминокислот из гемокапилляра к рибосомам гранулярной эндоплазматической сети; 2 - синтез и сегрегация белков; 3 - переход белков в вакуоли комплекса Гольджи; 4 - отщепление от комплекса Гольджи пузырьков с секреторными продуктами; 5 - выход секрета из клетки
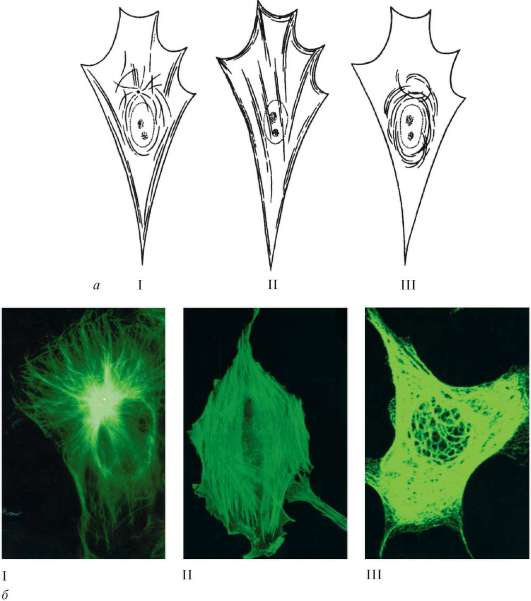
Рис. 4.16 Микрофиламенты и микротрубочки:
А - схема; б - микрофотографии (иммунофлюоресцентный анализ); бI - микротрубочки в культуре клеток фибробластов мыши (тубулин); бII - актиновые микрофиламенты в культуре клеток; бIII - промежуточные филаменты в культуре клеток эмбриональной почки свиньи
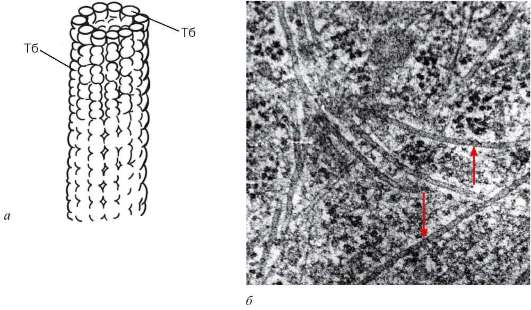
Рис. 4.17. Строение микротрубочек: а - Тб-субъединица, димер тубу-лина в составе микротрубочек; б - микротрубочки в цитоплазме клетки (стрелки)
Реснички и жгутики
Это специальные органеллы движения. В световом микроскопе эти структуры выглядят как тонкие выросты клетки. В основании реснички (fla-gellum) в цитоплазме видны хорошо красящиеся мелкие гранулы - базальные тельца. Длина ресничек составляет 5-10 мкм, а длина жгутиков может достигать 150 мкм (рис. 4.19). Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 300 нм. Этот вырост от основания до самой его верхушки покрыт плазматической мембраной. Внутри выроста расположена аксонема («осевая нить») - сложная структура, состоящая в основном из микротрубочек. Проксимальная часть реснички (базальное тельце) погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 200 нм). Базальное тельце по своей структуре очень сходно с центриолью. Оно также состоит из 9 триплетов микротрубочек. Часто в основании реснички лежит пара базальных телец, располагающихся под прямым углом друг к другу, подобно диплосоме.
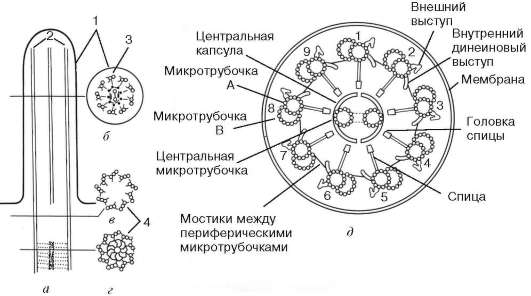
Рис. 4.19. Общее строение реснички:
А - продольный срез; б - поперечный срез тела реснички; в, г - срезы базального тельца. 1 - плазматическая мембрана; 2 - микротрубочки; 3 - дуплеты микротрубочек (А и В); 4 - триплеты микротрубочек базального тельца; д - схема поперечного среза реснички
Включения
Включения цитоплазмы -необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клеток.
Различают - трофические, секреторные, экскреторные и пигментные включения. К трофическим включениям - относятся капельки нейтральных жиров, которые могут накапливаться в гиалоплазме. В случае недостатка субстратов для жизнедеятельности клетки эти капельки могут постепенно исчезать, включаясь в обменные процессы. Другим видом включений резервного характера является гликоген - полисахарид, откладывающийся также в гиалоплазме (рис. 4.20). Отложение запасных белковых гранул обычно связано с активностью эндоплазматической сети. Так, запасы белка вителлина в яйцеклетках амфибии накапливаются в вакуолях эндоплазма-тической сети.
Секреторные включения - обычно округлые образования различных размеров, содержащие биологически активные вещества, образующиеся в клетках в процессе синтетической деятельности.
Экскреторные включения не содержат каких-либо ферментов или других активных веществ. Обычно это продукты метаболизма, подлежащие удалению из клетки.
Пигментные включения могут быть экзогенными (каротин, пылевые частицы, красители и др.) и эндогенными (гемоглобин, гемосидерин, билирубин, меланин, липофусцин). Наличие их в клетках может изменять цвет ткани и органа временно или постоянно. Нередко пигментация ткани служит одним из диагностических признаков некоторых заболеваний человека или характеризует возрастные изменения тканей и др.
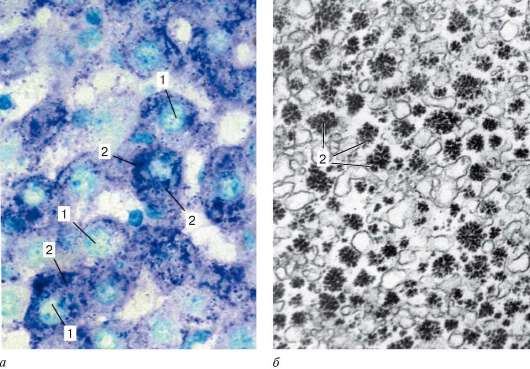
Включения гликогена в клетках печени:
Ядро
Ядро (nucleus) клетки -структура, обеспечивающая хранение и реализацию наследственной (генетической) информации, регуляцию синтеза белков.
Главными структурами, определяющими эти свойства, являются хромосомы, в ДНК которых содержится вся генетическая информация клеток. Хромосомы могут находиться в двух структурно-функциональных состояниях. В неделящихся, интерфазных клетках они находятся в различной степени деконденсации, или в рабочем состоянии, и представляют собой хроматин ядер интерфазных клеток. При делении клеток хроматин максимально уплотняется, конденсируется и образует собственно митотическую хромосому. Интерфазные хромосомы (хроматин) и митотические хромосомы представляют собой в химическом отношении идентичные образования.
Хроматин
При наблюдении живых или фиксированных клеток внутри ядра выявляются зоны плотного вещества, которые хорошо воспринимают разные красители, особенно основные. Благодаря такой способности хорошо окрашиваться этот компонент ядра получил название «хроматин» (от греч. chroma - цвет, краска). Такими же свойствами хроматина обладают и хромосомы, которые отчетливо видны как плотные окрашивающиеся тельца во время митотического деления клеток. В состав хроматина входит ДНК в комплексе с белками. В неделящихся (интерфазных) клетках хроматин, выявляемый в световом микроскопе, может более или менее равномерно заполнять объем ядра или же располагаться отдельными глыбками. Это связано с тем, что в интерфазном состоянии хромосомы теряют свою компактную форму, разрыхляются, или декон-денсируются. Степень такой деконденсации хромосом может быть различной. Зоны полной деконденсации хромосом и их участков морфологи называют эухроматином (euchromatinum). При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина, называемого гетерохроматином (heterochro-matinum). Степень деконденсации хромосомного материала - хроматина в интерфазе отражает функциональное состояние ядра клетки. Чем больший объем ядра занимает эухроматин, тем интенсивнее в нем протекают синтетические процессы.

Рис. 4.22. Ультрамикроскопическое строение ядра интерфазной клетки: 1 - ядерная оболочка (наружная и внутренняя мембраны, перинуклеарное пространство); 2 - комплекс ядерной поры; 3 - гетерохроматин (конденсированный хроматин); 4 - эухроматин (диффузный хроматин); 5 - ядрышко (гранулярная и фибриллярная части); 6 - межхроматиновые гранулы РНК; 7 - перихроматиновые гранулы; 8 – кариоплазма
В интерфазе фибриллы хроматина образуют петли. Эти петли собраны в розетки, где основания нескольких петель связаны друг с другом негистоно-выми белками ядерного матрикса. Такие петлевые группы (петлевые домены) при падении активности хроматина могут конденсироваться, уплотняться, образуя хромомеры, или хромоцентры, интерфазных ядер. Хромомеры выявляются также в составе митотических хромосом. Хромомеры тесно располагаются друг за другом и образуют новый фибриллярный уровень компактизации - хромонему. Последняя, далее конденсируясь, формирует основу хроматиды (хромосомы).
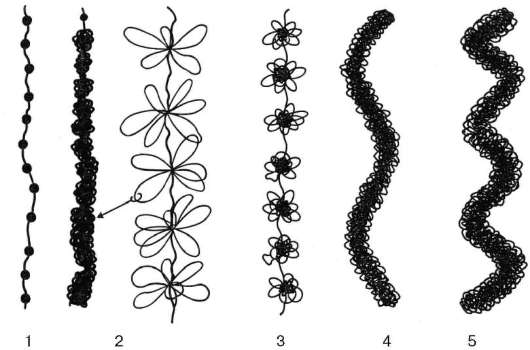
Схема различных уровней компактизации хроматина:
Хромонема; 5 – хроматида
Ядрышко
Практически во всех живых клетках эукариотических организмов в ядре видно одноили несколько обычно округлой формы телец величиной 1-5 мкм, сильно преломляющих свет, - это ядрышко, или нуклеола (nucleolus). К общим свойствам ядрышка относится способность хорошо окрашиваться различными красителями, особенно основными. Такая базофилия определяется тем, что ядрышки богаты РНК. Ядрышко - самая плотная структура ядра - является участком хромосомы, одним из ее локусов с наиболее высокой концентрацией и активностью синтеза РНК в интерфазе. Оно не является самостоятельной структурой или органеллой. Образование ядрышек и их число связаны с активностью и числом определенных участков хромосом - ядрышковых организаторов, которые расположены большей частью в зонах вторичных перетяжек; количество ядрышек в клетках данного типа может изменяться за счет слияния ядрышек или за счет изменения числа хромосом с ядрышковыми организаторами. ДНК ядрышкового организатора представлена множественными (несколько сотен) копиями генов рРНК: на каждом из этих генов синтезируется высокомолекулярный предшественник РНК, который превращается в более короткие молекулы РНК, входящие в состав субъединиц рибосомы.
Схему участия ядрышек в синтезе цитоплазматических белков можно представить следующим образом: на ДНК ядрышкового организатора образуется предшественник рРНК, который в зоне ядрышка одевается белком, здесь происходит сборка рибонуклеопротеидных частиц - субъединиц
рибосом; субъединицы, выходя из ядрышка в цитоплазму, организуются в рибосомы и участвуют в процессе синтеза белка.
Ядрышко неоднородно по своему строению: в световом микроскопе можно видеть его тонковолокнистую организацию. В электронном микроскопе выявляются две части: гранулярная и фибриллярная (см. рис. 4.22, б)
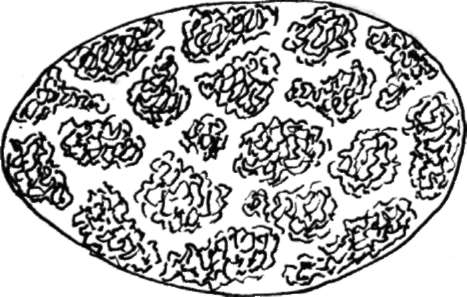
Рис. 4.25. Хромосомные территории в интерфазном ядре
Ядерная оболочка
Ядерная оболочка (tegmentum nucleare), или кариолемма, состоит из внешней ядерной мембраны (m. nuclearis externa) и внутренней мембраны оболочки (m. nucle-aris interna), разделенных перинуклеарным пространством (рис. 4.26). Ядерная оболочка содержит многочисленные ядерные поры (pori nucleares).
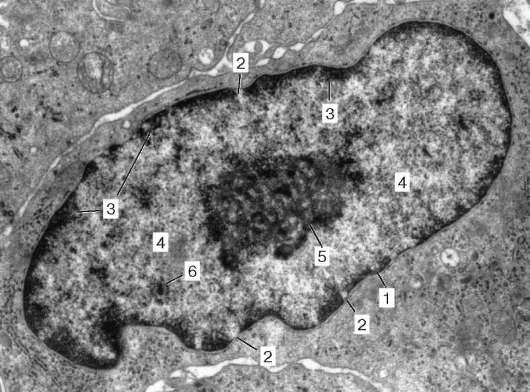
Рис. 4.26. Строение ядра интерфазной клетки:
Оболочка ядра (наружная и внутренняя мембраны, перинуклеарное пространство); 2 - комплекс ядерной поры; 3 - гетерохроматин; 4 - эухроматин; 5 - ядрышко; 6 - межхроматиновые гранулы РНК. Электронная микрофотография, увеличение 12 000
А - некроз; б - апоптоз.
Пояснения в тексте
ГИБЕЛЬ КЛЕТОК
ЦИТОЛОГИЯ. КЛЕТКА И НЕКЛЕТОЧНЫЕ СТРУКТУРЫ
Клетка -Основой строения эукариотических организмов является клетка (cellula) - наименьшая единица живого.
Эукариотические, собственно ядерные организмы — основная масса животных и растений, за исключением бактерий и сине-зеленых водорослей, не имеющих оформленного ядра — прокариотических организмов.






