

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...

Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...
Топ:
Оценка эффективности инструментов коммуникационной политики: Внешние коммуникации - обмен информацией между организацией и её внешней средой...
Устройство и оснащение процедурного кабинета: Решающая роль в обеспечении правильного лечения пациентов отводится процедурной медсестре...
Характеристика АТП и сварочно-жестяницкого участка: Транспорт в настоящее время является одной из важнейших отраслей народного...
Интересное:
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Искусственное повышение поверхности территории: Варианты искусственного повышения поверхности территории необходимо выбирать на основе анализа следующих характеристик защищаемой территории...
Берегоукрепление оползневых склонов: На прибрежных склонах основной причиной развития оползневых процессов является подмыв водами рек естественных склонов...
Дисциплины:
|
из
5.00
|
Заказать работу |
Содержание книги
Поиск на нашем сайте
|
|
|
|
ВВЕДЕНИЕ
Первичный океан, в котором зародилась жизнь (примем за основу идею самозарождения жизни на Земле), представлял собой довольно стабильную среду с постоянным химическим составом, температурой, давлением и освещением. Класс рыб на определенном этапе эволюции животного мира стал господствующим. Однако по мере остывания Земли, формирования суши, современной атмосферы часть рыб эволюционировала в наземных обитателей.
Позвоночные вырвались из зависимости от водной среды как среды обитания. С точки зрения эволюции Вселенной класс рыб остался в невыгодном положении и обречен на вымирание. По расчетам физиков, наше Солнце остынет через 8-10 млрд лет, а Земля превратится в такую же холодную планету, как Луна, Венера, Марс, непригодную для той формы органической жизни, которая сформировалась на Земле. Поскольку при остывании Земли уже при 0 њС вода перейдет в твердое состояние, то первыми от него пострадают рыбы. Та форма наземной жизни, которая обладает разумом, будет иметь шанс на выживание или за счет создания искусственной среды обитания, или переселения на другие пригодные для обитания космические тела.
Водная среда определила морфофункциональные особенности класса рыб. Наиболее специфичны у них органы дыхания. У рыб они достаточно эффективны, однако эволюция, не вполне полагаясь на жабры, вооружает водные организмы и другими специфическими инструментами извлечения кислорода из воды (плавательный пузырь, лабиринт, наджаберный орган, кожа и др.). В результате эффективность газообмена у рыб достигает 90 %.
Жизненно важное значение для рыб имеет осморегуляция. В пресной воде существует опасность нарушения гомеостаза за счет проникновения в организм избыточных количеств воды, а в соленой воде гомеостаз подвергается потрясениям противоположного характера - возникает опасность потери воды. В механизме поддержания осмотического давления участвуют многие системы органов, однако основная нагрузка приходится на почки и жабры рыб.
Рыбы очень чутко реагируют на изменения внешней среды; для водных животных адекватная и своевременная реакция на изменения среды обитания - вопрос жизни и смерти. Высокую чувствительность и реактивность к факторам; внешней среды обеспечивают хорошо развитый рецепторный аппарат и сенсорные системы рыб.
Рыбы воспринимают более широкий спектр стимулов из внешней среды по сравнению с высшими позвоночными животными.
Следует отметить важность появления у рыб сложного зрительного анализатора. Развитие его центральных механизмов создало предпосылки для эволюции головного мозга у наземных животных, усложнения их поведения, развития высшей нервной деятельности.
Изучение поведения рыб, у которых отмечается широкое разнообразие стереотипов врожденного одиночного, парного и группового поведения, позволяет лучше понять сложное поведение высших позвоночных животных, разработать оптимальные приемы управления поведением домашних животных. Анализ развития поведения в онтогенезе и механизма научения рыб в процессе жизненного опыта дает возможность специалистам научно обосновать процесс управления высшими позвоночными, а коррекцию неправильного поведения сделать более эффективной.
Специфична и система крови рыб. У рыб особые органы гемопоэза. У этих животных в широких пределах изменяются физико-химические свойства крови, для них характерен клеточный полиморфизм. Рыбы имеют один круг кровообращения, что предполагает и морфофункциональные особенности сердца.
Пищеварительная система рыб проявляет чудеса эволюционной пластичности. Пищеварительная трубка видоспецифична, ее строение изменяется непредсказуемо даже в пределах одного семейства. Причем наличие или отсутствие того или иного органа пищеварения не отражает эволюционного положения вида. Так, в семействе карповых известны безжелудочные мирные виды (карп, сазан, карась) и хищники с ампулообразным расширением пищеварительной трубки (голавль, жерех).
Спектр питания рыб очень широк, поэтому по типу питания рыб делят на несколько групп: от хищников до макрофитофагов, детритофагов и паразитов.
Не следует заблуждаться относительно происхождения целлюлозолитической активности кишки рыб, питающихся детритом и растительностью. Ее источник - симбиотические микробы, населяющие средний и каудальный участки кишки. Хитиназная активность желудочно-кишечного тракта также имеет экзогенное происхождение: это или автолиз, или результат микробной деятельности.
Морфофункциональная пластичность пищеварительной системы рыб служит предпосылкой для разработки рационов с включением различных кормовых средств.
В основе большого видового разнообразия класса рыб (около 25000 видов только костистых рыб), их широкого географического расселения и многочисленности популяций лежит необычайно высокая плодовитость. У некоторых видов рыб самки выметывают за нерестовый сезон сотни миллионов икринок. Однако незащищенность икры, вероятностный характер оплодотворения и уязвимость молоди на ранних этапах онтогенеза приводят к тому, что эффективность воспроизводства рыбы составляет всего несколько процентов. У живородящих рыб плодовитость ниже, но выживаемость молоди выше. При искусственном воспроизводстве эффективность размножения возрастает тысячекратно.
У рыб эндокринная регуляция тесно связана с нервной, контакты с которой прослеживаются на трех уровнях: клеточном, органном и организменном.
Стресс-реакции у рыб имеют тот же генерализованный ответ, что и описанный для млекопитающих Г. Селье. Однако постадийная стресс-реактивность рыб весьма специфична.
Рыбы имеют хорошо развитый механизм специфической и неспецифической иммунной защиты. У них те же механизмы реализации иммунитета, что и у высших позвоночных: клеточный ответ осуществляется за счет фагоцитоза, пиноцитоза, цитотоксического влияния - инструментов, которыми обладают у рыб не только все формы лейкоцитов, но и эритроциты, а в особых случаях даже тромбоциты. Гуморальный ответ осуществляется посредством лизоцима, интерферона, пропердина, а также гамма-глобулинов. Ветеринарная иммунология в рыбоводстве находится в зачаточном состоянии, однако имеет очень хорошие перспективы развития.
Большое значение для понимания физиологических потребностей рыбы и комплексной оценки ее состояния имеют этологические наблюдения. Отклонение от нормального поведения рыбы должно настораживать исследователя. "Неправильное" поведение свидетельствует или о несоответствии среды обитания потребностям рыбы, или об изменении констант внутренней среды организма рыб. При искусственном рыборазведении и в первом, и во втором случае потребуется вмешательство специалиста. Характер вмешательства должен быть обоснован с позиции физиологии Рыбы, понимания ее потребностей и возникающих мотиваций поведения.
Таким образом, рыба как объект хозяйственной деятельности человека требует к себе особого внимания и прежде всего, более глубокого понимания физиологии поведения
обмена веществ, размножения, зоогигиенических требований к среде обитания. Знание физиологических особенностей рыбы создает основу для разработки современных технологических систем производства рыбной продукции или для выращивания декоративных рыб, в которых полностью ревизуется генетический потенциал объекта разведения.
К сожалению, авторы современной учебной литературы, обходят стороной особенности физиологии рыб, а зачастую просто вводят читателя в заблуждение в этой области физиологии. Специальные учебники и учебные пособия для студентов по физиологии рыб единичны и издавались давно. В последние годы в специальной литературе появились интересные материалы по морфофункциональным особенностям рыбы, которые ввиду языковых и иных барьеров недоступны для студентов и специалистов. Многолетний опыт исследовательской работы с рыбами, преподавания основ физиологии рыб студентам и аспирантам Московской сельскохозяйственной академии имени К. А. Тимирязева, а также систематический анализ научной литературы по физиологии рыбы побудили автора к написанию этого учебника.
Автор надеется, что учебник будет полезен для студентов, аспирантов, а также специалистов, занятых рыборазведением.
Автор готов к сотрудничеству со всеми заинтересованными лицами. Все замечания от читателей будут приняты с благодарностью.
|
|
|
|
|
|
|
|
Глава 4. КОЖНЫЙ ПОКРОВ
Кожа рыб выполняет ряд важных функций. Располагаясь на границе внешней и внутренней среды организма, она выступает фактором неспецифического иммунитета, защищая рыбу от внешних воздействий. Одновременно, отделяя организм рыбы от окружающей ее жидкой среды с растворенными в ней химическими веществами, кожа рыб является эффективным гомеостатирующим механизмом.
Для многих видов рыб кожный покров - основное средство защиты от неблагоприятных факторов. Кожная слизь характеризуется высокой бактерицидной активностью. Благодаря большому содержанию тканевой тромбокиназы и кининов (фактор свертывания крови) в кожной слизи обеспечивается высокая скорость свертывания крови при травмировании рыбы.
В роли средств защиты тела рыбы от механических повреждений выступает жесткая чешуя, покрывающая кожу большинства рыб. Чешуя (плакоидная) акул, скатов, хрящевых рыб по прочности и строению напоминает кости. Поэтому некоторые исследователи называют ее кожными зубами.
Кожа многих морских рыб снабжена железами клетки, вырабатывающими яды (рис. 4.1).
Секретом этих желез покрыты колючки плавников и шипы на жаберных крышках. Хищники стараются избегать контактов с такими рыбами.
Яды многих ядовитых рыб, относящиеся к группе нейротоксинов, оказывают быстрый отравляющий эффект. Например, тетродотоксин - яд наиболее опасных рыб (сем. иглобрюхие, или рыбы-собаки) - в 10 раз токсичнее яда кураре. Он смертелен для человека.
Не менее опасна и бородавчатка страшная (семейство бородавчатки), распространенная в Индийском и Тихом океанах. В спинном плавнике этой рыбы находится 13 колючек с ядовитыми железами.
Рана, нанесенная этой рыбой, болезненна и вызывает паралич конечности человека. При попадании в рану большого количества яда у человека развиваются одышка, сердечная недостаточность, судороги, расстройства психики, а в тяжелых случаях может наступить смерть.
Наиболее ядовит морской дракончик (сем. морские дракончики) - обитатель Черного моря. У него ядовитые железы расположены у основания колючек спинного плавника и в основании шипов жаберных крышек. Сильными ядами снабжены плавники морского ерша (скорпены) и глубоководных морских окуней (себастосы). Последние являются объектом промысла и потому вызывают тяжелые заболевания рук у рыбаков.
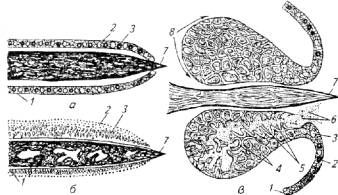
Рис. 4.1. Ядовитые железы рыб:
а- ядовитые железы (одноклеточные) плавниковой колючки морского ерша: б - ядовитые железы хвостового шипа ската-хвостокола; в -ядовитые железы жаберной крышки морского дракончика: 1 - эпидермис; 2 - слизистые клетки; 3 - железистые клетки; 4 - опорные клетки; 5 - проток; 6 - ядовитый секрет; 7- шип; 8 - ядовитая железа
|
|
Пигментные клетки кожи придают рыбе определенную окраску, что важно для ее маскировки. В общении индивидуумов окраска рыбы имеет, кроме того, информационное значение. Наиболее распространена серебристая окраска рыб, которую обеспечивает пигмент гуанин. Пигментные клетки с кристаллами гуанина обычны для большинства рыб. Они расположены не только в дерме кожного покрова, но и на чешуе. Поэтому чешуя и имеет серебристый цвет. В промышленности рыбья чешуя используется для производства пата, из которого получают искусственный жемчуг.
В коже рыб находится много нервных окончаний. Поэтому ее можно рассматривать как орган рецепции (механической, термической, химической, электрической).
Для многих рыб кожа является органом газообмена (кожное дыхание). Слизистый покров кожи защищает некоторых рыб от высыхания на воздухе (угри, сомы), обеспечивая поглощение кислорода из воздуха.
У рыб кожа выполняет также довольно специфическую опорную - функцию. На внутренней стороне кожи рыб закрепляйся мышечные волокна скелетной мускулатуры, составляющие миомеры. Таким образом, она выступает как опорный элемент в составе опорно-двигательного аппарата.

Рис. 4.2. Строение кожи рыб (обобщение):
1 - эпидермис; 2 - дерма; 3 - подкожная клетчатка; 4 - бокаловидные клетки; 5 - оруглые клетки; 6- колбовидные клетки; 7 - пигментные клетки.
Наконец, физические свойства кожи таковы, что создают особые условия для эффективного перемещения в плотной водной среде. Скорости плавания таких рыб, как мерлин, тунцы, скумбрии и др., настолько высоки, а энергетические затраты рыб при этом настолько низки, что ставят в тупик физиков, пы-тающихся описать движение рыб классическими законами для неживых физических тел. Тело рыбы при движении в воде оказывает меньшее гидродинамическое сопротивление по сравнению с неживым телом той же формы, массы и плотности.
Большую роль в снижении гидродинамического сопротивления играет кожа, способная понижать коэффициент трения воды и тела, а также особым образом создавать обтекающие тело рыбы водные потоки.
Кожа рыб состоит из двух слоев: наружного, или эпидермиса, и нижнего, или соединительнотканного, - дермы (рис. 4.2).
Между ними выделяют базальную мембрану. У большинства высших рыб эпидермальный слой тонкий и мягкий. Он пронизан чувствительными нервными окончаниями, но не содержит кровеносных сосудов. В дерме просматриваются как нервные элементы, так и капиллярная сеть.
§15. ЭПИДЕРМИС
Эпидермис кожи рыб представлен многослойным эпителием, состоящим из 2-15 рядов клеток. Клетки верхнего слоя имеют плоскую форму. Часть клеток верхнего ряда ороговевает и отторгается. Нижний (ростковый) слой представлен одним рядом цилиндрических клеток, которые, в свою очередь, происхо-дят от призматических клеток базальной мембраны. Средний слой эпидермиса состоит из нескольких рядов клеток, форма которых изменяется от цилиндрической до плоской.
В эпидермисе могут быть заложены слизистые клетки трех типов. У базальной мембраны располагаются колбовидные клетки, чуть выше, в среднем слое, просматриваются округлые клетки, а у поверхности, в слое плоских клеток, находятся бокаловидные слизистые клетки. Однако все три типа клеток одновременно присутствуют лишь у некоторых видов рыб, т. е. в виде исключения.
У быстроплавающих рыб (тунцы, мерлин) в эпидермисе представлены только округлые клетки, выделяющие серозный секрет. У пелагических рыб, плавающих со средними скоростями, помимо округлых клеток имеются бокаловидные клетки. У донных и других малоподвижных рыб можно обнаружить все три типа клеток.
Секреты слизистых клеток отличаются по своему химическому составу и имеют различное предназначение. Так, слизь кожи тунца не такая густая, как у сома, но очень богата белком. Секрет колбовидных клеток обычно не содержится в кожной слизи, так как колбовидные клетки лежат в глубине эпидермиса и не имеют протоков. Их секрет попадает на поверхность тела лишь при травмировании кожного покрова. Однако секрет колбовидных клеток обладает сильным биологическим эффектом. Его называют "веществом испуга" - настолько сильно его воздействие на окружающих травмированную особь рыб.
Низкомолекулярные соединения из кожи хищных рыб (кайромоны) несут информацию о присутствии в водоеме хищника. Причем этот сигнал воспринимается как мирными, так и хищными рыбами. В свою очередь, мирные рыбы выделяют через кожу феромон тревоги, который обладает межвидовой активностью. Однако феромон своего вида производит более выраженное воздействие на сородичей по сравнению с феромоном других даже генетически близких видов рыб. Феромон тревоги выделяется рыбами под влиянием не только биотических факторов (хищник), но и гипоксии, изменения рН среды, а также в результате антропогенного воздействия (например, контрольный облов пруда, механизированное выкашивание водной растительности и др.).
Было установлено, что чувствительность половозрелых рыб к феромону тревоги чрезвычайно вели-ка и составляет для большинства исследованных видов 0,1-0,01 мкг сырой кожи на 1 л воды. Для некоторых видов рыб пороговая концентрация была на 3-4 порядка ниже.
Секрет бокаловидных клеток имеет густую консистенцию и содержит сравнительно большое коли-чество мукополисахаридов, гликопротеидов и производных холестерина.
Слизь как продукт кожного эпидермиса играет важную роль в Жизни рыб. Кожа активных пловцов (например, тунцов и скумбрий) вырабатывает меньше слизи, чем кожа менее подвижных рыб (сом, налим, вьюн). Количество слизи положительно коррелирует с уровнем развития кожного дыхания рыб.
Роль слизи как средства снижения коэффициента трения в литературе сильно преувеличена: активные пловцы - мерлин, тунцы, акулы - имеют жесткую и шершавую кожу с малым количеством слизи.
Химический состав слизи у рыб разного филогенетического положения и из разных экологических ниш различается незначительно. Плотность составляет 1,01 - 1,03 г/см3, pH 5,0-5,4. Значительно большее влияние на состав слизи оказывают факторы среды. Так, в зависимости от степени возбуждения (стрессиро-ванности) рыбы ее кожная слизь содержит 0,25-3,0 г/л белка, 2-5 мг/д гемоглобина, 0,1-0,4 ммоль/л кетонов, а также углеводный комплекс сложного состава (гексозы, мукополисахариды, сиаловые кислоты) и нуклеиновые кислоты.
При стрессах в слизи обнаруживаются эритроциты (до 500 в 1 мм3) и лейкоциты.
§16. ДЕРМА
Дерма (кутис) состоит из трех слоев: тонкого верхнего (соединительнотканного), толстого среднего сетчатого слоя коллагеновых и эластиновых волокон и тонкого базального из высоких призматических клеток, дающих начало двум верхним слоям.
У активных пелагических рыб дерма хорошо развита. Толщина ее в участках тела, обеспечивающих интенсивное движение (например, на хвостовом стебле акулы), сильно увеличена. Средний слой дермы у активных пловцов может быть представлен несколькими рядами прочных коллагеновых волокон, которые между собой связываются еще и поперечными волокнами.
У медленноплавающих литоральных и донных рыб дерма рыхлая или вообще слаборазвитая.
Под дермой находится рыхлый слой соединительной ткани с жировыми включениями (подкожная клетчатка). У быстроплавающих рыб на участках тела, обеспечивающих плавание (например, хвостовом стебле), подкожная клетчатка отсутствует. В этих местах к дерме прикрепляются мышечные волокна. У других рыб (чаще всего медлительных) подкожная клетчатка хорошо развита и включает много жира (например, у зубатки или нототении).
Как известно, для рыб характерна определенная окраска, что справедливо прежде всего для тропических рыб. Последние отличаются как цветовой гаммой, так и яркостью раскраски. Цвет рыбы обусловлен наличием в дерме пигментных клеток - хроматофоров и лейкофоров (рис. 4.3).

Рис. 4.3. Пигментные клетки кожи рыб:
1 - меланофоры, 2- ксантофоры и эритрофоры; 3- гуанофоры
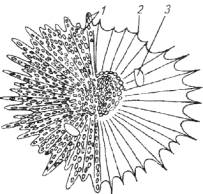
Рис. 4.4. Пигментная клетка рыбы:
1 - зерна пигмента; 2 - фибриллы; 3 - клеточное ядро
В этих клетках обнаруживается, по крайней мере, четыре типа пигмента. Окраска кожи рыбы формируется как результат сочетания нескольких пигментов. По поверхности тела пигментные клетки рассредоточены неравномерно. В результате на теле образуются узоры различного цвета, яркости и формы (полосы, пятна, кольца и др.).
Большинство видов рыб, обитающих в водоемах средней полосы, окрашено скромно. Спинка у этих рыб пигментирована в темный цвет, что делает их незаметными с берега. Бока и брюшко имеют серебристую окраску, что позволяет рыбе слиться с бликующей на солнце поверхностью воды. Рыбы, обитающие в зарослях (окунь, щука), имеют маскировочные серо-зеленые поперечные полосы. Донные рыбы (сом, ротан, пескарь) принимают окраску дна водоема, зачастую копируя при этом не только цвет, но и рисунок грунта.
Меланофоры содержат зерна пигмента черного цвета. Эритрофоры содержат красный пигмент, ксантофоры - желтый, лейкофоры (гуанофоры) - кристаллы гуанина (см. рис. 4.3).
Пигментная клетка отличается особым строением. Цитоплазма в ней неоднородна. Верхний слой (эктоплазма) неподвижен, так как пронизан особыми жесткими органеллами - радиальными фибриллами (рис. 4.4).
Внутренний слой цитоплазмы пигментной клетки (киноплазма) подвижен и содержит зерна пигмента. Киноплазма обладает способностью концентрироваться в центре клетки или растекаться по всей клетке. При этом происходит или концентрация пигмента в единую каплю (клетка светлеет), или распределение пигмента по всей площади клетки (клетка окрашивается).
§17. ОКРАСКА ТЕЛА
Окраска рыбы может иметь различное биологическое значение. Она позволяет рыбе оставаться незамеченной при нападении на жертву (полосы у щуки или окуня скрывают рыбу в зарослях подводной растительности). В ряде случаев стратегия окрашивания прямо противоположна - яркой окраской тела рыба пытается напугать врага или пищевого (полового) конкурента.
Брачные наряды имеют другую цель - привлечь внимание полового партнера.
Изменение окраски кожи происходит под влиянием нервной и гуморальной систем. Быстрое "перекрашивание" обеспечивается нервной регуляцией. Центры пигментации тела у рыб лежат спинном мозге, у разных видов в разных сегментах. Так, пигментно-моторные волокна спинного мозга у карповых переходят в симпатическую цепочку на уровне 12-18-го позвонков, у камбалы - на уровне 5-6-го позвонков. Отсюда они распространяются в краниальном и каудальном направлениях. У карповых перерезка симпатической цепочки у 12-го позвонка вызывает потемнение передней части тела, а перерезка за 18-м позвонком приводит к потемнению задней части тела.
Гуморальная регуляция пигментации тела обеспечивает более поздние изменения окраски. Парентеральное введение гормонов (адреналин, аденокортикотропный гормон, гистамин) приводит к изменению пигментации через несколько десятков минут.
Решающее значение в запуске реакций пигментации у рыб имеет зрительная афферентация. Например, камбала принимает окраску тела в зависимости от того, на грунте какого цвета находится ее голова. При этом на теле воспроизводится не только тон, но и рисунок (полосы, пятна).
Реакция пигментации тела запускается быстро: уже через 5 мин камбала окрашивается в соответствии с тоном и рисунком грунта (рис. 4.5). Однако завершается пигментация в течение 1-5 ч. Процесс по-темнения занимает меньше времени, чем процесс посветления.
Существуют и межвидовые различия в скорости пигментации и депигментации тела.
Так, у трески потемнение кожи заканчивается за несколько минут, в то время как у камбалы на это уходит несколько часов. Морской петух, бойцовая рыбка и скорпена при раздражении изменяют свою окраску за несколько секунд. Это свидетельствует о том, что у разных видов рыб соотношение нервного и гуморального звеньев в регуляции пигментации тела различно.
У быстро изменяющих окраску своего тела рыб активность хроматофоров в большей мере подконтрольна нервной системе. Ослепленные рыбы утрачивают способность изменять окраску. Очевидно, что брачный наряд рыб (горбуша, бычки, колюшки) возникает вследствие гормональных перестроек.

Рис. 4.5. Зависимость окраски тела камбалы от цвета грунта, на котором находится ее голова
§18. ЧЕШУЯ
Подавляющее большинство пластинчатожаберных и костистых рыб имеют чешую. Однако ее количество на коже и размер отдельных чешуек сильно различаются у разных видов рыб. У скатов, бычков и сомов чешуя редуцирована. У угрей чешуя очень мелкая и практически невидима невооруженным глазом.
Б водоемах разных широт обитают рыбы с очень крупной чешуей. Например, у старых особей зеркального карпа и индийского усача диаметр отдельных чешуек достигает нескольких сантиметров.
Характер чешуйчатого покрова рыб определяется их этолого-экологическим положением. Активные пловцы литоральной зоны любого водоема имеют хорошо развитую крупную чешую, обеспечивающую хорошую обтекаемость тела. Чешуя препятствует образованию складок кожи при движении, особым образом организует обтекающие потоки и защищает мышечные волокна, прикрепленные к дерме, а также внутрен-ние органы от давления воды. В особых случаях чешуя защищает рыбу от зубов хищника.
Химический анализ чешуи рыб показывает, что она на 50 % состоит из органического вещества, представленного в основном соединительной тканью. Минеральную часть образует главным образом фосфат кальция (около 40 %). В меньших количествах в чешуе обнаружен карбонат кальция и натрия, а также фосфат магния. Довольно разнообразен и микроминеральный состав чешуи.
Среди рыб распространена чешуя трех типов, различающихся по химическому составу и форме: плакоидная, ганоидная, костная.
Плакоидная чешуя является самой древней в филогенетическом плане. Она свойственна ископаемым рыбам, а также современным акулам и скатам. Отдельные чешуйки имеют форму ромбической пластинки с шипом, который выступает из кожи наружу. Минеральная часть представлена дентином, который пропитывает соединительнотканную основу чешуи. Шип плакоидной чешуи отличается очень высокой прочностью, так как снаружи покрыт особой эмалью - витродентином. Плакоидная чешуйка имеет полость, заполненную рыхлой соединительной тканью с кровеносными сосудами и нервными окончаниями (рис. 4.6, I). У некоторых видов хрящевых рыб плакоидная чешуя видоизменена. Например, у морской лисицы на поверхности тела имеются крупные бляшки. Колючки скатов также являются преобразованными плакоидными чешуйками.
Ганоидная чешуя появляется на более поздней стадии филогенеза рыб. Ее можно наблюдать у многоперов, осетровых (на хвосте), кистеперых рыб. Ганоидные чешуйки имеют вид ромбических толстых пластин, соединяющихся между собой посредством особых сочленений (рис. 4.6, II). Поэтому ганоидная чешуя, как правило, формирует характерный плотный панцирь, кожные кости или щитки на коже рыб. Прочность ганоидной пластинке придает в верхней части ганоин, а в нижней - костное вещество.

Рис. 4.6. Чешуя современных рыб:
I- плакоидная чешуя; II-ганоидная чешуя; III- костная чешуя; а - сельдь; 6- лещ; в- окунь
Костная чешуя современных рыб отличается тем, что в ней присутствует лишь костное вещество. В зависимости от формы костная чешуя подразделяется на циклоидную и ктеноидную (рис. 4.6, III). Циклоидная чешуя (карповые, сельдевые рыбы) имеет форму округлой и гладкой пластинки. Ктеноидная (окуневые рыбы) отличается от циклоидной чешуи наличием мелких шипиков по заднему краю.
Циклоидная и ктеноидная чешуя очень похожи не только морфологически. Они имеют идентичный химический состав. Один вид рыб может иметь оба типа чешуи. Так, у камбалы самцы имеют ктеноидную, а самки - циклоидную чешую.
Характерной особенностью костной чешуи является наличие на ней концентрических колец.
Кольца образуются в результате неравномерного роста рыбы в целом и прироста чешуи в частности (рис. 4.7).
Исследования показали, что между ростом тела и ростом чешуи рыбы существует прямолинейная зависимость, которая описывается уравнением
Ln=(Vn/V)L,
где Ln - ожидаемая длина рыбы в возрасте п; Vn - расстояние от центра чешуи до годового кольца в возрасте n; V - длина чешуи от центра до края; L - длина рыбы.
Рыба растет всю жизнь. Однако рост ее тела и отдельных его частей идет неравномерно на протяжении как всей жизни, так и каждого года ее жизни. В летнее время рыба активно питается и потому растет быстрее. Па период зимовки рост рыбы замедляется или вообще прекращается.
При активизации обменных процессов в организме рыбы происходит и ускоренный рост чешуи, что сопровождается образованием широкого темного кольца. Периоды замедленного роста отмечаются на чешуе в виде светлых полос и колец (рис. 4.8).
Закладка годовых колец у разных рыб приходится на разное время года. Известно, например, что у молоди азовского судака закладка годового кольца приходится на весну, у половозрелых особей - на вторую половину лета. У молоди леща годовое кольцо закладывается весной, у рыб старших возрастов - осенью. Следовательно, закладка годового кольца определяется не только температурными колебаниями и интенсивностью питания, но и наследственными факторами.
У неполовозрелых и половозрелых рыб линейный рост тела по времени не совпадает. Молодь начинает расти с весны, в то время как половозрелые рыбы первую половину вегетационного периода накапливают питательные вещества.
Образование годового кольца есть результат взаимодействия факторов внешней среды с обменными процессами организма рыбы посредством нейрогуморальных регулирующих механизмов. Это соображение подтверждает и то, что годовые кольца отмечены у рыб в тропическом поясе, где сезонные колебания температуры воды и колебания доступности пищи отсутствуют. Тем не менее многие жизненные функции тропических рыб цикличны. Для них характерны циркадные, лунные и годичные циклы.

Рис. 4.7. Изменение размеров тела и чешуи в процессе роста рыбы (прямолинейная зависимость)

Рис. 4.8. Годичные кольца на чешуе рыб:
1-вобла; 2 -пресноводная колюшка; 3-треска; 4-лосось; а, б, в,-годичные; г, д -добавочные кольца; е - край чешуи
§19. ВОЗРАСТ РЫБ
Возраст рыб можно определить по количеству концентрических колец на чешуе.
А. Левенгук (1684) первым обратил внимание на соответствие числа колец на чешуе количеству прожитых рыбой лет. Методика подсчета колец довольно проста. Для определения возраста обычно используется чешуя с участка у основания первого спинного плавника.
Чешую промывают в растворе нашатыря, протирают, зажимают между двумя предметными стеклами и просматривают под лупой или при малом увеличении микроскопа. (Мелкую чешую перед просмотром целесообразно смазать глицерином.)
Часто между годовыми кольцами просматриваются добавочные кольца, происхождение которых связывают с нерестом (лососи, сельди) или изменением интенсивности питания (карповые рыбы). Добавочные кольца выражены не по всей длине, а лишь с какой-то одной стороны чешуи.
Следует признать, что не все виды рыб имеют чешую с ясно различимыми годовыми кольцами. Поэтому для уточнения результата подсчета количества годовых колец используют годовые кольца костей. Для этой цели подходят различные кости: у окуней - жаберная крышка, у осетровых - лучи грудных плавников, У судака и сома - лучи спинного или анального плавника (рис. 4.9).
Возраст многих морских рыб Удобнее определять по годовым кольцам на отолитах (камбала, нототения, треска). В этих случаях производят поперечный распил костей или делают шлифы. Годовые кольца рассматривают под лупой (шлифы лучей - под микроскопом), при необходимости для большей четкости препараты смазывают глицерином или смачивают водой.
Трактовка полученных результатов может быть затруднительной или неоднозначной. Поэтому для рыб обитающих в

Рис. 4.9. Годовые кольца на спиле луча рыб, грудного плавника севрюги
водоемах средних и высоких широт, приняты определенная терминология и специальные обозначения для характеристики возраста рыб по годовым кольцам.
Количество колец на чешуе (костях, лучах, отолитах) и календарный возраст рыб увязаны следующим образом:
| Число колец | Обозначение | Термин |
| Нет | 0+ | Сеголеток |
| Одно | Годовик | |
| Одно | 1+ | Двухлеток |
| Два | Двухгодовик | |
| Два | 2+ | Трехлеток |
| Три | Трехгодовик | |
| и т.д. |
Движение следует рассматривать как важнейшее свойство всего живого. На субклеточном уровне наравне с раздражимостью клетки оно является отличительным признаком жизни. Однако движение в микромире характерно как для растений, так и для животных. Движение многоклеточных структур при помощи специализированных клеток и тканей (мышечных) является отличительной особенностью животных организмов. Появление способности к движению обозначило новый этап эволюции. Возникновение поперечнополосатой мускулатуры у животных позволило им завоевать всю биосферу.
В эволюционном плане именно движение послужило поводом для возникновения нервной системы в животном мире. По мере усложнения двигательной активности животных происходило усложнение их нервной системы. В пределах любого класса как беспозвоночных, так и позвоночных животных уровень развития нервной системы отражает степень сложности двигательной активности. У позвоночных животных уровень развития опорно-двигательного аппарата коррелирует со сложностью строения нервной системы и определяет сложность поведенческих реакций, обеспечивающих питание, расселение, размножение, освоение новых территорий, внутри- и межвидовых отношений.
С усложнением движения связан эволюционный прогресс. Животные, по каким-то причинам утратившие подвижность (например, паразиты), представляют собой тупиковую ветвь эволюционного дерева. Все крупнейшие эволюционные изменения связаны с совершенствованием двигательной функции. Такие ароморфозы, как выход из водной среды на сушу, легочное дыхание, внутреннее оплодотворение, млекопитание новорожденных, обусловлены прогрессивным развитием функции движения.
Движение позволяет животным эффективно приспосабливаться к изменению внешней среды, т. е. сохранить свою жизнеспособность, сберечь популяцию и вид в целом. В основе приспособляемости организма лежат два двигательных процесса: уклонение (защита) от неблагоприятного фактора или изменение самого себя. В первом случае животное использует опорно-двигательный аппарат (уклоняется от опасности, находит пищу, строит убежище). Во втором случае животное адаптирует свой организм: усиливает кровообращение, повышает частоту дыхания, усиливает (сокращает) теплоотдачу и т. д. Однако в обоих случаях исполнительными органами выступают структуры, включающие сократительные элементы.
§20. ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Морфологическую основу движения образует опорно-двигательный аппарат. Собственно движителем выступают мышцы. Именно в мышце происходит трансформация химической энергии АТФ в механическую энергию. Однако мышце для сокращения и производства движения нужна точка опоры. Такими точками опоры для многочисленных мышц рыбы выступают кости скелета. Скелет выполняет и формообразующую функцию (рис. 5.1).
Строение скелета рыб (рис. 5.2). По многообразию форм тела рыб можно судить и о сложности строения их скелета (рис. 5.2). Особенностью рыб является то, что многие из них имеют как традиционный для всех позвоночных животных внутренний, так и наружный скелет. Последний можно рассматривать как признак эволюционного застоя. У костистых рыб наружный скелет представляет только чешуя. Однако у осетровых рыб наружный скелет довольно хорошо развит. Собственно чешуя у них присутствует лишь на хвостовом стебле, а туловищная часть и голова несут на себе костные образования - жучки, бляшки, колючки и шипы, доставшиеся современным рыбам от их предков - панцирных рыб.
У рыб требования к жесткости и прочности костей ниже, чем у наземных позвоночных. Следует отметить, что и относительная масса костей у рыб в 2 раза меньше. Размеры скелета костистых рыб меняются пропорционально массе тела. Эта зависимость может быть описана уравнением регрессии:
Мск=0,033Мтела1,03,
где Мск- масса скелета, г; Мтела, - масса тела, г.
Меньшая масса костей для водных животных очень важна, Имея большой удельный вес, костная ткань существенно влияет на плавучесть тела водных животных. Поэтому даже вторично водные животные (китообразные) в процессе своей адаптаци
|
|
|
Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...

Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций...

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰)...

Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!

