Лиганды
Рецепторы (старое наименование) | СС хемокины | СХС хемокины| Предоминантная экспрессия/распределение по тканям | Патогены, использующие данный рецептор для инфицирования | Локализация на хромосоме
СС-рецепторы CCR1 (СС CKR1) | МIР-1а, b RANTES МСР-3 | — | Моноциты, Т-клетки | — | Зр21
CCR2A (МСР-1Ra) | МСР-1 | — | Т-клетки, базофилы, моноциты | — | Зр21
CCR2B (MCP-1Ra) | МСР-1, 2, 3 | — | — | ВИЧ-1 (NSI) | —
CCR3 (CKR3) | Эотаксин, RANTES, МСР-2, 3, 4 | — | Базофилы, эозинофилы, микроглиальные клетки, и, возможно, моноциты; небольшая экспрессия в дендрических клетках или Т-лимфоцитах периферической крови | ВИЧ-1 (NSI) | Зр21
CCR4 | TARC | — | Базофилы, Т-клетки | — | Зр24
CCR5 (СС CKR5) | MlP-la, P RANTES | — | Моноциты, дендрические клетки, макроглиальные клетки, Т-клетки | ВИЧ-1 (NSI), ВИЧ-2 | Зр21
СХС рецепторы CXCR1 (IL-8 RA) | — | IL-8 | Нейтрофилы, НК-клетки | — | 2q35
CXCR2 (IL-8 RA) | IL-8, MGSA, gro-альфа, NAP-2, IP-10, ENA-78, Mig | — | — | — | —
CXCR3 | — | IP-10, Mig | Активированные Т-клетки | — | —
CXCR4 (фузин, LESTR, HUMSTR) | — | SDF-1 | Очень распространен: CD4+- и CD4– клетки, моноциты, макрофаги, дендрические клетки, В-клетки; другие ткани, например, мозг, легкие, селезенка | ВИЧ-1 (SI), ВИЧ-2 | 2q21
СС/СХС рецептор DARC (Duffy-антиген) | RANTES, MCP-1, TARC и т. д. | IL-8, MGSA, gro-альфа и т. д. | Эндотелиальные клетки, эритроциты | Plasmodium vivax | 1
Другие STRL33 | Неопределены | Неопределенны | Лимфоидная ткань и активированные Т-клетки | — | —
HCMV-US28 | MlP-l альфа, бета, RANTES | — | Фибробласты, инфицированные CMV | ВИЧ-1 | Нет
ChemRl | Неопределены | Неопределенны | Т-лимфоциты, полиморфно-ядерные клетки | — | Зр21-24
CMKBRL1 | Неопределены | Неопределенны | Нейтрофилы, моноциты, мозг, печень, легкие, скелетные мышцы | — | Зр21
TER1 | То же | То же | Тимус, селезенка | — | Зр21
V28 | То же | То же | Нервная и лимфоидная ткани | — | Зр21
D2S201E | То же | То же | Распространен широко, включая клетки гемопоэтического происхождения | — | Зр21
BLR1 | То же | То же | В-лимфоциты | — | —
ЕВН | То же | То же | То же | — | —
GPR 1,2,5 | — | — | — | Неопределены | —
Сокращения: BLR1 рецептор 1 лимфомы Буркитта; CMKBRL1 — хемокин подобный бета-рецептор 1; DARC — duffy-антиген/рецептор для хемокинов; ЕВН — рецептор, индуцируемый вирусом Эпштейна — Барра; ENA78 — полученный из эпителия нейтрофилактивирующий пептид-78; GPR — G-хемокин, присоединяющий рецептор; gro — связанный с ростом продукт гена; HCMV — цитомегаловирус человека; HUMSTR — рецептор человеческого сывороточного трансмембранного сегмента; IL — интерлейкин; IP-10 — интерферон-гамма индуцибельный 10 кд белок; LESTR — сегмент лейкоцитэспрессируемого трансмембранного рецептора; МСР — белок хемотаксиса лимфоцитов; Mig — монокины, индуцируемые гамма-интерфероном; MIP — макрофаговый воспалительный белок; NSI — не индуцирующие синтиций вирусы; NAP2 — нейтрофилактивирующий белок 2; RANTES — регулируемые при активации, нормальные Т-клеточные экспрессированные и секретированные хемокины; SDF-1 — фактор 1, производный из клеточной стромы; STRL33 — рецептор, включающий 7 трансмембранных доменов, полученный из 33-го лимфоцитарного клона; TARC — хемокины, регулируемые тимусом и активацией [McNicholl J. еt al., 1997]
Первыми ключ к разгадке роли хемокиновых рецепторов в развитии ВИЧ-инфекции подобрали исследователи знаменитой лаборатории д-ра Галло, обнаружившие, что высокий уровень хемокинов ингибирует репликацию ВИЧ в условиях in vitro [Cocchi F. et al, 1995]. Вскоре публикации работ о роли хемокиновых рецепторов в развитии ВИЧ-инфекции и, в частности, рецепторов CXCR4 и CCR5, приобрели взрывной характер. Установлена связь этих рецепторов с возбудителями инфекционных болезней (рис. 49).
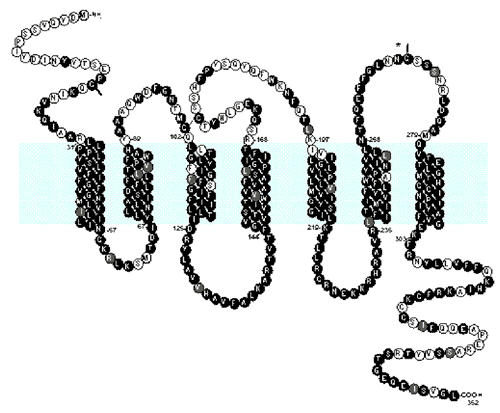
Рис. 49. Предполагаемая структура и аминокислотная последовательность CCR5. Обычно этот рецептор представлен тремя внутриклеточными, тремя внеклеточными и семью трансмембранными доменами. Горизонтальная окрашенная полоса соответствует клеточной мембране. Аминокислотные остатки, соответствующие CCR1b, окрашены в черный цвет. Светлого цвета консервативные структуры [McNicholl J. еt al., 1997].
Точный механизм, благодаря которому ВИЧ проникает в клетку посредством CXCR4 и CCR5, и роль рецептора CD4 в этом механизме, неизвестны. Предполагается, что пенетрация вируса осуществляется посредством взаимодействия VЗ-петли и других участков его наружного оболочечного белка gp120 и внеклеточных доменов CCR5- и ССR2b-рецепторов, и может включать многоэтапное взаимодействие с CD4 и другими клеточными поверхностными компонентами. Хемокины могут блокировать проникновение вируса посредством блокирования рецептора, снижения его афинности или ингибирования последующих этапов процесса пенетрации [Oravecz Т. et al., 1996].
Делеция в 32 bp образуется в участке повторяющегося мотива гена CCR5. В результате сдвигается рамка считывания в кодирующей рецепторный белок нуклеотидной последовательности, и синтезируемая дефектная форма рецептора утрачивает три из 7 трансмембранных доменов, 2 из 3 наружных петель и внутриклеточный сигнальный домен [McNicholl J. et al., 1997] (рис. 50, 51 и 52).

Рис. 50. Предполагаемая структура и аминокислотная последовательность мутантной формы рецептора CCR5. Мутантный белок утрачивает, по крайней мере, три трансмембранных сегмента CCR5 из 7 трансмембранных доменов, 2 из 3 наружных петель и внутриклеточный сигнальный домен — в результате ВИЧ не может «узнать» клетку-мишень [McNicholl J. еt al., 1997].
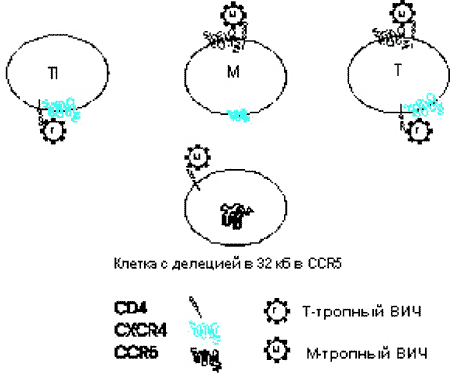
Рис. 51. Хемокиновые рецепторы и клеточный тропизм к ВИЧ. На схеме показаны три типа клеток — пассированные Т-клеточные линии (Т1), моноцит/макрофаг (М) и циркулирующие Т-клетки (Т). Т-клеточные линии экспрессируют CXCR4, но не CCR5; макрофаги и циркулирующие в периферической крови Т-клетки экспрессируют оба рецептора, хотя количество CXCR4 у них меньше чем на макрофагах. М-тропность у ВИЧ вызвана способностью белковых структур его оболочки связываться с CCR5 и проникать в оба типа макрофагов, а также в циркулирующие в крови Т-клетки. Т-тропный ВИЧ преимущественно связывается с CXCR4 и проникает в Т-клетки или Т-клеточные линии. После связывания с рецепторами хемокинов и рецептором CD4, вирус сплавляется с клеточной мембраной. Клетки с 32 bр делецией в гене CCR5 не экспрессируют кодируемый им белок и, хотя М-тропный ВИЧ может связываться с CD4, он не может проникнуть в клетку. Если клетки экспрессируют CXCR4, они могут быть инфицированы Т-тропным вирусом [McNicholl J. еt al., 1997].

Рис. 52. Участок CCR5 гена и аминокислотная последовательность с делецией в 32 bр. Повышенную частоту встречаемости 32 bр делеций гена CCR5 в человеческой популяции, можно рассматривать как своеобразный генетический шрам, оставленный прошлыми эпидемиями СПИДа. Интересно, что африканцы, которые непосредственно контактируют с природным резервуаром возбудителя СПИДа среди африканских приматов, оказывается, менее знакомы с ним, чем европейцы. Рисунок J. McNicholl и соавт., 1997.
Является ли резистентность к ВИЧ у людей, гомозиготных по мутации в рецепторе CCR5, абсолютной? К сожалению, нет.
Другие рецепторы, например, CCR3 и CCR2b также являются кофакторами проникновения в клетки отдельных серотипов ВИЧ. Т. е. Люди с генотипом CCR5 ▲32/▲32 лишь относительно резистентны к ВИЧ. Однако эта относительная устойчивость может быть довольно значительной. По крайней мере, описан один пациент, у которого болезнь не развилась даже после переливания инфицированной крови [O'Brien Т. et al., 1997]. У лиц с одной копией мутантного гена развитие болезни затягивается на два года по сравнению с лицами, гомозиготными по гену дикого типа [McNicholl J. et al., 1997].
Распределение в популяциях риска мутации гена рецептора CCR5, имеет четко выраженный этнический и расовый характер. По данным различных авторов у кавказцев гомозиготные лица в популяции могут составлять 1 % при 10–20 % гетерозиготных [McNicholl J. et al., 1997]. Этот ген встречается у 20 % людей белой расы [Garred P., 1998]. У афроамериканцев в популяции до 6 % гетерозигот, у испанцев до 7 %, у азиатов менее 1 %. Среди африканцев, таиландцев, японцев и корейцев практически не встречаются даже гетерозиготы по делеции в гене CCR5 [McNicholl J. et al., 1997]. Наличие большого количество таких гомозигот и гетерозигот среди европейцев свидетельствует о том, что они уже сталкивались с эпидемиями СПИДа. Они и привели к «отбору» генотипов, относительно устойчивых к ВИЧ. Следовательно, повышенную частоту встречаемости 32 bр делеций гена CCR5 в популяции можно рассматривать как своеобразный генетический шрам, оставленный прошлыми эпидемиями СПИДа. Остаются не ясными причины, благодаря которым африканцы, контактировавшие с природным резервуаром ВИЧ среди африканских приматов, оказывается, менее с ним знакомы, чем европейцы. Этот вопрос мы рассмотрим в главе 3.
Из генов других хемокиновых рецепторов, мутации в которых снижают восприимчивость человека к ВИЧ, исследователи отмечают производные CCR2. Замена в 64 позиции гена валина изолейцином приводит к тому же эффекту, что и делеция 32 bр в гене CCR5. Эффект обоих мутаций на прогрессирование ВИЧ-инфекции является аддитивным [McNicholl J., 1998].
Роль МНС в устойчивости к ВИЧ. Было показано, что на передачу ВИЧ-1 от инфицированной женщины ее потомству во время беременности и родов влияют аллели DRB1 МНС класса I младенца. Сорок шесть ВИЧ-инфицированных и 63 сероревертирующих младенцев, родившихся с пассивноприобретенными анти-ВИЧ антителами, и не ставших инфицированными, типировали, используя автоматический метод ДНК-диагностики. Одна или более DR-13-аллелей, включая DRB1 5*01301, 1302 и 1303 были обнаружены у 15,2 % детей, ставших ВИЧ-инфицированными и у 31,7 % сероревертирующих младенцев (р = 0,048); эта ассоциация зависела от этнического происхождения ребенка. После аналогичного исследования в отношении других аллельных ассоциаций, выяснилось, что только DR2-аллель DRB1 5*0 1501 ассоциировалась с серореверсией у относящихся к кавказской расе детей. Среди этих младенцев аллель DRB1 5*0 03З011 положительно ассоциировалась с ВИЧ-инфекцией [Singh N. et al., 1997].
По предварительным данным Multi Center AIDS Cohort Study, CCR5 и МНС генотипы влияют на устойчивость к ВИЧ независимо друг от друга. По оценкам специалистов Центра влияние МНС на этот процесс значительно большее, чем у мутаций хемокиновых рецепторов [Kaslow R. et al., 1997].
Таким образом, чувствительность к ВИЧ у различных популяций людей сильно варьирует. Наиболее чувствительными к вирусу являются азиатские и негроидные этнические группы. В тоже время устойчивость к инфицированию вирусом других рас носит относительный характер. Это предполагает невозможность торможения эпидемии СПИДа в результате формирования устойчивых к ВИЧ популяций.
Вирус герпеса saimiri. H. saimiri — онкогенный лимфотропный гамма-герпесвирус — трансформирует человеческие и обезьяньи Т-клетки в условиях in vitro и вызывает лимфомы и лейкемию у различных видов приматов Нового Света. Открытая рамка считывания генома H. saimiri кодирует сильно гликозилированный белок, который выделяется и связывается с гетеродимерными HLA-DR-молекулами МНС класса II. Эти результаты показывают, что открытая рамка считывания может функционировать как иммуномодулятор, способный приводить к иммунопатологии при инфекции H. saimiri [Singh et al., 1997].
Вирус Денге. Исследования L. Zeng и соавт. (1996) позволяют предполагать, что реакция Т-клеток на вирус Денге ограничивается аллелью HLA-DR15.
Вирус гепатита В. Поскольку HLA-антигены класса I содержат сходные с вирусными пептиды, они могут быть важными мишенями для иммуноопосредованного гепатоцитолиза CD8+ CTL при гепатите В. Davenport и соавт. (1995) показали, что HLA-DR13 ассоциируется с резистентностью к гепатиту В.
Вирус гепатита С. У больных, инфицированных вирусом гепатита С, прогноз может быть совершенно разным: у половины пациентов наблюдается хроническая болезнь печени, тогда как у другой половины признаков гистологических изменений, связанных с повреждением печени, не обнаруживают. Иммунные реакции хозяина могут играть важную роль при таких разных результатах. К. Кигоkochi и соавт. (1996) идентифицировали цитотоксический Т-клеточный эпитоп, представленный HLA-A2 в NSS-области вируса гепатита С. В исследовании, проведенном G. Реапо с соавт. (1994), было установлено, что антиген HLA-DR5 выступает как защитный фактор против тяжелого исхода болезни, вызванной вирусом гепатита С.
Вирус Эпштейна — Барра. Предположение о влиянии реакции CTL на эволюцию вируса Эпштейна — Барра было впервые высказано на основании обнаружения того факта, что вирусные изоляты от высокопозитивных на HLA-A11 людей азиатского происхождения, специфически мутировались в двух иммунодоминант-АИ-ограниченных CTL-эпитопах. Кроме того, В35.01-ограниченные реакции CTL у белых доноров воспроизводимо картируются до одного пептидного эпитопа. Однако большинство изолятов вируса Эпштейна — Барра от людей, живущих там, где преобладал аллель В35.01 (в Гамбии), либо сохраняли последовательность эпитопа CTL, либо несли мутацию, которая сохраняла антигенность. Изменения, ведущие к пониженной антигенности, были обнаружены лишь в небольшом количестве случаев [Singh N. et al., 1997]. Два эпитопа, специфичных для вируса Эпштейна — Барра CTL, идентифицированы общим аллелем HLA-B7 [Hill и соавт., 1995].
Хантавирус Puumala. В то время как в США хантавирусы проявляют себя преимущественно легочными симптомами, в Северной Европе их основным патологическим проявлением является почечный синдром. Хотя большинство инфекций, вызываемых этими вирусами, вероятно имеют субклинические или мягкие формы, у 10 % людей болезнь может прогрессировать и приводить к шоку, 5 % больных нуждаются в диализе, а некоторые могут даже погибнуть. У тех, кто перенес болезнь в форме почечного синдрома, повреждения почек впоследствии могут проявиться хронической гипертензией. Так как хантавирусы являются вариабельными и обычно передаются «роями», исследователями было предположено, что такие факторы людей, как HLA-гены, сильно влияют на спектр клинических проявлений болезни [McNicholl J., 1998].
Действительно, это было показано в конкретных случаях. Лица, которые экспрессировали гены HLA-B8, переносили болезнь более тяжело. У них болезнь проявлялась пониженным кровяным давлением, повышенным содержанием креатинина в крови. Одновременно наблюдалось большее количество вируса в моче и крови, определяемого с помощью ПЦР. У лиц с HLA-B27 болезнь протекала в мягкой форме. Почти все больные с гаплотипом HLA-A1-B8-DR3, у которых прогрессивно развивался шок, экспрессировали аллель TNF2. Аллель была связана с повышенной продукцией фактора некроза опухолей [Wilson A. et al., 1997].
Возбудитель болезни Лайма. B. burgdorferi — человеческий патоген, передаваемый переносчиками. Вызывает мультисистемную болезнь, поражающую кожу, нервную систему, сердце или суставы. Артриты являются главным поздним проявлением болезни. Хотя все клинические проявления болезни Лайма поддаются лечению антибиотиками, приблизительно 10 % больных с артритами имеют хронические воспаления суставов, длящиеся месяцы и даже годы после антибиотикотерапии. У таких пациентов после проведения курса антибиотикотерапии, ПЦР-тесты на ДНК B. burgdorferi в жидкости суставных сумок были отрицательными. Этот факт позволяет предположить, что воспаление сустава может продолжаться даже после того, как спирохеты будут из него удалены, поэтому оно носит не инфекционный характер [McNicholl J., 1998].
Исследование аллелей HLA класса II показало, что аллель HLA-DRB1*0401 связана с хроническими артритами и отсутствием у организма реакции на антибиотикотерапию. Эта аллель связана также с повышенным риском развития тяжелых ревматоидных артритов. При исследовании антительных ответов у пациентов, перенесших болезнь Лайма, реакции иммуноглобулина G (IgG) на поверхностный наружный белок A (OspA) и OspB спирохеты часто обнаруживались в самом начале длительно текущих артритов. После проведения курса лечения у пациентов с HLA-DR4 и антительными ответами на OspA и OspB, артриты продолжались значительно дольше, чем в тех случаях, когда ответы на данные белки отсутствовали [Kalish R. et al., 1993]. Это вызвано тем, что лица, переболевшие болезнью Лайма и страдающие устойчивыми к лечению артритами, обычно имеют Т-клетки, которые взаимодействуют со многими эпитопами OspA, в то время как у лиц, хорошо отвечающих на лечение, они обычно отсутствуют. Т. е. Т-клеточный ответ на OspA у больных с артритами, устойчивыми к лечению, может перекрестно распространяться и на собственный антиген в суставной сумке. И тогда ответ на этот собственный антиген может продолжать вызывать воспаление сустава в течение месяцев или даже лет после удаления из него спирохет [McNicholl J., 1998].
Возбудитель туберкулеза. Устойчивость к туберкулезу (ТВ) сильно варьирует у этнических групп.
Врожденная недостаточность функции фагоцитоза. На хромосоме 2q35 человека был идентифицирован ген NRAMP1, гомологичный мышиному Bcg-гену, который придает мышам резистентность к бациллам Кальмета и Герена и к возбудителю лепры (палочка Гансена). Хотя все четыре мутантных типа этого гена были связаны с ТВ, два из них обнаружены у больных туберкулезом в особенно большом количестве. Отдельные аллели гена с необыкновенно высокой частотой преобладают в восточно-африканской популяции, но редко встречаются в европейских. Это наблюдение может частично объяснить более высокую, в сравнении с другими этническими группами, чувствительность африканцев к туберкулезу. Хотя физиологическая функция гена NRAMP1 точно не определена, она может иметь отношение к функции фагоцитолиза [McNicholl J., 1998].
Врожденная недостаточность системы интерферона. Мутация в гамма-интерфероновом рецепторе 1 (IFNгаммаRl) приводит к диссеминации малопатогенных микобактерий. Ген IFNгаммаRl расположен в регионе хромосомы 6q22q23. Мутация в гене приводит к отсутствию экспрессии рецептора на поверхности клеток. Мутация часто обнаруживается в семьях больных туберкулезом [Newport M. et al., 1996].
Врожденная низкая усвояемость витамина D. В случаях ТВ, но не в контролях, также наблюдается низкая встречаемость генотипа, гомозиготного по мутантному рецептору витамина D. Одновременно этот генотип ассоциируется с повышенным риском развития остеопорозов [McNicholl J., 1998].
Связь восприимчивости с МНС. Выявлена связь между легочной формой ТВ и специфичностью HLA. HLA-DR2 ассоциируется с развитием множественно-палочковидных форм как ТВ, так и лепры. Молекулярное субтипирование DR2 показало, что значительную часть аллеля у больных, но не контролей, составляли DRB1 5+ 01501 и DRB1 5+ 01502. Частота этих молекулярных подтипов DR2 у больных не изменялась, позволяя предположить, что вся молекула DR2 или тесно связанный с ней ген(ы) способны определять восприимчивость больного к легочному ТВ и, в частности, к ТВ резистентному к лекарственным препаратам [Singh N. et al., 1997].
Mycobacteriumleprae. Роль в развитии лепры врожденной недостаточности функции фагоцитоза, связанной с мутантной аллелью гена NRAMP1 на хромосоме человека 2q35, рассмотрена выше (см «Возбудитель туберкулеза»).
HLA-аллели модулируют иммунную реакцию, определяющую форму лепры. На одном полюсе клинического спектра лепры находятся больные с мультибациллярной (multibacillary) лепроматозной лепрой (LL), которые являются иммунологически толерантными к антигенам М. leprae. На другом — больные с туберкулоидной лепрой (ТТ) с малым количеством палочек (paucibacillary), обнаруживающие хорошую клеточно-опосредованную иммунную реакцию. Гуморальный иммунитет присутствует по всему спектру, но, по-видимому, не обеспечивает защиты. Между двумя полюсами находятся больные с промежуточными признаками, как при погранично-лепроматозной, пограничной, и погранично-туберкулоидной формах. Было зарегистрировано увеличение частоты HLA-DR2 и — DQ1 у больных с LL и HLA-DR3 у больных с ТТ. Эти антигены можно еще подразделить на аллели, определенные по их аминокислотным последовательностям. Одна аминокислотная замена может привести к появлению аллелей с различными иммунологическими свойствами. Аллель DRB1 5*01501 больше ассоциировался с больными LL, чем с больными ТТ (р<0,00001). Кроме того, аллели DQB1 5*00601 обнаруживали значительно чаще у больных с LL, чем у контрольных больных (р<0,00001); DQA1 5*00103 чаще встречался в группе с LL, чем в группе с туберкулезной лепрой; и частота DQA1 5*00102 избирательно повышалась у больных с погранично-лепроматозной лепрой. Однако частота аллелей DRB1 5*00701, DQB1 5*00201 и DQA1 5*00201 снижалась у больных LL по сравнению с больными ТТ и контрольными пациентами, а частота DQB1 5*00503 избирательно уменьшалась у больных ТТ, позволяя предположить, что эти аллели могли модулировать иммунную реакцию, которая определяет форму лепры, развивающуюся у каждого больного [Rani R. et al., 1993].
Малярия. Естественная устойчивость к паразитарным болезням носит более сложный характер, чем к бактериальным и вирусным. Известно три вида естественной устойчивости к малярии [Шувалова Е.П., 1973]:
1. Абсолютная устойчивость, например, невосприимчивость обезьян резус к P. vivax.
2. Фазовая устойчивость, относящаяся к определенным стадиям паразита. Тканевые клетки могут быть восприимчивы, а кровь иммунна, как, например, в случае устойчивости обезьяны шимпанзе к P. ovale.
3. Неполная устойчивость, заключающаяся в том, что развиваться могут все стадии паразита, но их размножение ограничено (например, устойчивость у шимпанзе к P. vivax).
Малярийные плазмодии — это первый, ставший известным, фактор отбора, оставивший «генетические шрамы» в отдельных человеческих популяциях.
Роль генетических дефектов гемоглобина в устойчивости к малярии. Duffy-мутация является единственной защитой хозяина против P. vivax, не приводящая к другим серьезным последствиям (Duffy-антиген — рецептор хемокинов; см. табл. 27). Такие гемопоэтические модификации, как талассемия, G6PD-дефицитность и гемоглобин S, препятствуют развитию плазмодия, но у гомозиготных людей они сами становятся причиной развития серьёзных болезней [Lederberg J., 1997].
Наиболее изученным генетическим дефектом гемоглобина, препятствующим развитию малярийного плазмодия, является замена глутамина на валин в шестом положении (бета-цепи гемоглобина; гемоглобин HbS). Указанное замещение серьезно отразилось на функциональных свойствах гемоглобина. HbS в восстановленной форме образует длинные ассоциированные цепи, его растворимость снижается на 50 % по сравнению с растворимостью окисленной формы. Эти особенности гемоглобина определяют укорочение жизни эритроцитов в связи со снижением их толерантности к гемолизу [Бышевский А.Ш., Тарсенов О.А., 1994]. У гомозиготных людей в детском возрасте серповидноклеточная анемия отличается более тяжелым течением и высокой летальностью. Она осложняется калькулезными холециститами, язвами конечностей (в области лодыжек), остеомиелитами, переломами костей, ретинопатией, гематурией, тромбозами сосудов головного мозга и другими тяжелыми клиническими симптомами. Однако у гетерозигот болезнь проявляется в менее тяжелой форме, но при этом люди с HbS формой гемоглобина не заболевают малярией. Даже трудно себе представить цену, уплаченную человеческими популяциями в эндемичных по малярии регионах Африки «демону Дарвина», за частоту аллеля серповидноклеточности в 20 % (рис. 53).

Рис. 53. Распространение серповидно-клеточной анемии в Африке. Малярийные плазмодии — это первый, ставший известным, фактор отбора, оставивший «генетические шрамы» в отдельных человеческих популяциях. В ряде районов Африки распространена наследственная болезнь — серповидно-клеточная анемия. Эритроциты больного человека принимают форму серпа и часто разрушаются, возникает тяжелое малокровие, в результате чего человек отстает в развитии и даже погибает. Казалось бы отбор должен в течение одного-двух поколений устранить этот летальный ген серповидно-клеточности. Но оказалось, что в полной мере эта болезнь проявляется только у гомозиготных по данному локусу людей. У гетерозигот анемия проявляется в менее выраженной форме, но вместе с тем они обладают невосприимчивостью к другой страшной и смертельной в этих местах болезни — малярии. Поэтому там, где есть малярия, естественный отбор поддерживает динамическое равновесие между концентрацией летального серповидно-клеточного аллеля и частотой «здорового» аллеля, или, как принято говорить у эволюционных биологов, поддерживает в популяции балансированный полиморфизм. Частота аллеля серповидно-клеточности: 1–15–20 %; 2–10–15 %; 3–5–10 %; 4 — менее 5 % [Медников Б.М., Яблоков А.В., 1987].
Роль комплекса HLAв устойчивости к малярии. Поскольку существуют значительные различия между экспонируемыми и неэкспонируемыми малярией людьми в частотах HLA-генов по локусам А и В, HLA-комплекс может защищать население на эндемичных по этой болезни территориях от паразитов, вызывающих малярию. Адекватные механизмы могут экспрессироваться HLA-ассоциированными генами, которые контролируют иммунологическую реактивность к антигенам малярии. Хорошо установлена связь между HLA-антигеном класса I, HLA-B53, и защитой от тяжелой формы малярии [Hill A. et al., 1992]. Эта связь может быть опосредованна ограниченными HLA класса I цитотоксическими Т-лимфоцитами на стадии пребывания паразита в печени. Среди иммунных к малярии африканцев НLА-В53-ограниченные CTL распознавали сохраненный неамерпептид из антигена, специфического для печеночной стадии, но ни один НLА-В53-ограниченный эпитоп не был идентифицирован в антигенах из других стадий развития паразита [Нill A. et al., 1992]. Эти данные демонстрируют возможную молекулярную основу для установления связи HLA с болезнью и поддерживают кандидатуру антигена, специфического для стадии печени, в качестве компонента вакцины против малярии [Singh N. et al., 1997].
Роль гена фактора некроза опухолей в клинических проявлениях малярии. Независимо от HLA-аллелей, аллель TNF2 гена фактора некроза опухолей альфа, показана как фактор, способствующий развитию церебральных форм малярии [McGuire W. et al., 1994]. От распространенного аллеля TNF1, TNF2 отличает более сильный транскрипционный активатор, способствующий увеличению экспрессии TNF-альфа. Видимо по этой причине у больных с церебральными формами малярии обычно находят повышенное содержание в крови TNF-альфа [Miller L., 1996]. Эта же аллель способствует более тяжелому течению инфекции, вызываемой хантавирусом Puumala (см. «Хантавирус Puumala»).
Роль гена внутриклеточного фактора адгезии 1 в клинических проявлениях малярии. Отдельные аллели гена ISAM-1 способствуют повышенной адгезии паразита к клеткам красной крови в эндотелии небольших сосудов, это сопровождается повышением чувствительности людей к церебральным формам малярии [Fernandez-Reyes D. et al., 1997].
Шистосоматоз. В исследованиях, проведенных с помощью сегрегационного анализа, показано, что среди населения Бразилии ген SM1 несет ответственность за 66 % случаев тяжелого течения болезни (рис. 54). Приблизительно 3 % бразильцев гомозиготны по этому гену и предрасположены к тяжелым формам течения болезни (АА-лица), 68 % гомозиготны по резистентности шистосоматозу (аа-лица), 29 % гетерозиготны и имеют промежуточный уровень резистентности к паразиту (Аа-лица).

Рис. 54. Распределение гена, определяющего чувствительность популяции к возбудителю инфекционной болезни. На схеме показана частота аллеля А гена SM1, предрасполагающего к развитию шистосоматоза, среди населения Бразилии [Abel L., Desseint A., 1998].
Ген SM1 картирован в регионе хромосомы 5q31-q33, ответственном за контроль дифференциации Т-лимфоцитов [Marquet S. et al., 1996]. В дальнейших исследованиях этот регион был связан и с локусом, отвечающим за синтез иммуноглобулина Е (IgE) и образование эозинофилов. В регуляции устойчивости человека к шистосоматозу ген SM1 участвует через регулирование лимфокинами популяций Th2. Резистентные гомозиготы усиливают Th0/2-ответ, в тоже время чувствительные гомозиготы проявляют в отношении паразита Th0/1-ответ [Abel L., Desseint A., 1998].
* * *
Восприимчивость к возбудителю инфекционной болезни человеческой популяции или этнической группы всегда носит полигенный характер и определяется частотами встречаемости генов, продукты экспрессии которых способны усилить инфекционный процесс. Частоты таких генов в популяциях варьируют. Поэтому в любой популяции наблюдается множество переходных вариантов патологии между типичными формами проявления инфекционной болезни.
Генетическая устойчивость к развитию микробной инфекции может включать устойчивость к возникновению инфекции, т. е. устойчивость в первоначальной стадии размножения микроорганизма в хозяине и устойчивость к дальнейшему развитию болезни, т. е. Удержание инфекции в состоянии латентности. Генетическая устойчивость к паразитарным болезням носит более сложный характер, чем к бактериальным и вирусным, и предполагает наличие фазовой устойчивости, относящейся к определенным стадиям развития паразита.
Генетическая устойчивость (предрасположенность) к отдельным возбудителям инфекционных болезней может реализовываться непосредственно через иммунную систему и минуя её. Среди факторов иммунной системы, специфически связанных с развитием отдельных инфекций, наиболее изучена роль факторов, кодируемых генами МНС, ассоциации с инфекциями отдельных аллелей которых имеют универсальный характер. Показана роль в развитии инфекционных болезней мутаций генов регулирующих функцию фагоцитолиза (например, NRAMP1 для возбудителей ТВ и лепры); регулирующих активность иммунной системы (например, врожденная недостаточность системы интерферона при ТВ); контролирующих дифференциацию Т-лимфоцитов (например, дефектный ген SM1 при шистосоматозе) и др.
Среди генетических факторов, специфически влияющих на чувствительность к возбудителям инфекций, но не реализующихся через иммунную систему, можно выделить следующие: мутации в рецепторах, нарушающих механизм лиганд-рецепторного взаимодействия возбудителя с клеткой-мишенью (например, Duffy-мутация повышает устойчивость эритроцитов к малярийным плазмодиям, мутации гена рецептора CCR5 снижает риск инфицирования ВИЧ); мутации в субстратах, метаболизируемых узкоспециализированным паразитом (например, генетические дефекты гемоглобина определяют устойчивость к плазмодиям, вызывающим малярию); мутации в генах рецепторов, нарушающих взаимодействие клетки с витамином D и др.
Устойчивость к инфицированию возбудителями инфекционных болезней у рас, этнических групп, отдельных популяций, всегда носит относительный характер. После каждой крупной эпидемии из-за элиминации отдельных генотипов, чувствительность популяции меняется в отношении нескольких возбудителей инфекционных болезней одновременно.






