В годы, последовавшие за клонированием гена per, было предложено несколько моделей, объясняющих то, как его белковый продукт PER может функционировать для обеспечения циркадных колебаний.
Была предложена модель “мембранного градиента”, в которой PER играл роль насоса для создания градиента, который спадал бы за счёт светочувствительных каналов при достижении порогового значения. В другой модели белок PER был представлен как протеогликан, который соединяет клетки, тем самым облегчая образование межклеточных связей через щелевые контакты.
Прорыв был наконец достигнут с подтверждением наличия PER-антител. Первым результатом стало определение 24-часового колебания концентрации PER-белка в нейронах головного мозга мух с максимальным значением в ночное время (Siwicki et al., 1988). Для иРНК, кодируемой геном per, также были выявлены суточные колебания концентраций в клетках мозга мух, что указало на зависимость цикличности белка PER от цикличности иРНК гена per. Интересно, что пик уровня иРНК гена per наблюдался ранней ночью, за несколько часов до пика концентрации белка PER (Hardin et al., 1990). Значим тот факт, что ген per с миссенс мутацией был не способен обеспечить циклические колебания концентрации иРНК гена, но белок PER дикого типа смог восстановить циклическую экспрессию иРНК. Исходя из этих наблюдений, родилась модель саморегулируемой отрицательной обратной связи, при которой накопление белка PER приводит к ослаблению экспрессии иРНК гена per (Hardin et al., 1990). Впоследствии было обнаружено, что белок PER относится к нуклеопротеинам и перемещается между ядром клетки и цитоплазмой с определённой заданной периодичностью, подтверждая идею о том, что он является регулятором транскрипции (Liu et al., 1992).
В последующем исследовании Янг обнаружил ген timeless, вспомогательный ген, влияющий на циркадные ритмы (Myers et al., 1995; Sehgal et al., 1995). В серии последовательных экспериментов лаборатория Янга выявила, что уровни иРНК гена timeless также обладают 24-часовым периодом колебаний, и что TIM может напрямую присоединяться к PER, влияя на его локализацию в ядре и концентрацию, блокируя распад PER (Gekakis et al., 1995; Sehgal et al., 1994; Vosshall et al., 1994). Важно отметить, что цикл экспрессии гена per был нарушен у мух с мутацией гена timeless, и наоборот, циркадные циклы в экспрессии гена timeless были расстроены у особей с мутантным геном per. (Sehgal et al., 1994, 1995). Эти результаты позволили окончательно обозначить концептуальные рамки TTFL как механизма обеспечения циркадных ритмов через ауторегуляцию генов биологических часов (Рис. 2А). В то же время пока не был окончательно ясен механизм транскрипции, что давало почву для существования различных теорий.
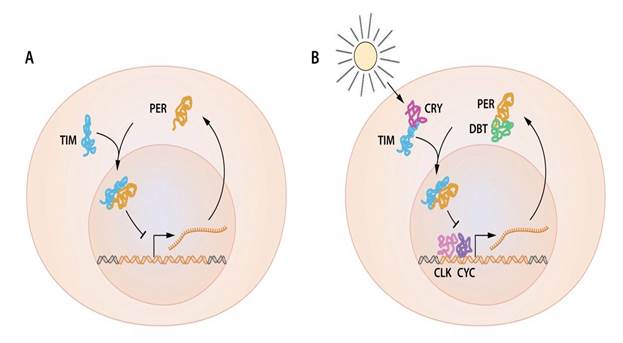
Рисунок 2 | Упрощенная схема регуляции гена по типу обратной связи.
а | И мРНК гена per, и белок PER подвержены ритмичным колебаниям, причем белок PER накапливается спустя несколько часов после пика мРНК гена per. Белок PER локализуется в ядре, и активность гена per колеблется вследствие того, что белок PER ингибирует его по типу обратной связи.
b | Вспомогательные белки необходимы при регуляции колебаний гена per. TIM белок, кодируемый геном timeless, также осциллирует и взаимодействует с белком PER. Это взаимодействие имеет критическую важность для накопления в ядре белка PER и репрессии гена per. DBT-белок кодируется геном doubletime. DBT представляет собой протеинкиназу, которая фосфорилирует PER, приводя к его разрушению. DBT-опосредованный распад белка PER обуславливает задержку между накоплением мРНК гена per и его продуктом. CLK и CYK, кодируемые часовыми генами и цикл-генами, являются двумя факторами транскрипции, которые активируют ген per.
Очевидно, что открытие самоподдерживающегося механизма TTFL является совершенно новой парадигмой. Однако, принцип, по которому активировалась транскрипция генов per и timeless, оставался неизвестным. Этот вопрос разрешился с обнаружением часового и цикличного генов (Allada et al., 1998; Rutila et al., 1998). Часовой ген был впервые идентифицирован у мышей Джозефом Такахаси (King et al., 1997). Взаимодействующие друг с другом продукты часового гена CLOCK (CLK) и CYCLE (CYC) содержат основные домены ”спираль-петля-спираль“ (bHLH) и связываются с определёнными участками соответствующих генов, тем самым положительно регулируя их транскрипцию. В последующих исследованиях было выяснено, что TIM и PER выступают как отрицательные регуляторы активности CLK, замыкая таким образом циркадную петлю обратной связи (Darlington et al., 1998).
Современные рабочие модели циркадных молекулярных часов чрезвычайно сложны и включают в себя множество дополнительных компонентов, способствующих стабильности и циркадной периодичности (Hardin, 2011). Важно отметить, что, поскольку реакции транскрипции и трансляции, как правило, являются быстрыми, то для совершения 24-часовых колебаний основной механизм транскрипционно-трансляционной обратной связи должен осуществляться с существенной задержкой. Это достигается обширным комплексом реакций, включающих в себя регулируемое фосфорилирование белков и распад компонентов TTFL, сборку белкового комплекса, ядерную транслокацию и другие посттрансляционные модификации (Hardin, 2011). Ключевые данные, проливающие свет на основополагающий механизм такой задержки, были получены после открытия Янгом гена doubletime, кодирующего протеинкиназу DOUBLETIME (DBT), которая, в свою очередь, фосфорилирует PER и увеличивает скорость его распада (Price et al., 1998). Добавочные белки объединяют сигналы извне, которые могут влиять на внутренние часы (рисунок 2B). К примеру, свет может активизировать белковый продукт криптохромного гена (CRY) и способствовать его связыванию с TIM, что приводит к протеасомному распаду последнего (Ceriani et al., 1999; Emery et al., 1998). С наступлением утра TIM распадается, благодаря чему PER может быть фосфорилирован DBT и впоследствии расщеплён.






