

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...

Своеобразие русской архитектуры: Основной материал – дерево – быстрота постройки, но недолговечность и необходимость деления...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Своеобразие русской архитектуры: Основной материал – дерево – быстрота постройки, но недолговечность и необходимость деления...
Топ:
Оснащения врачебно-сестринской бригады.
Генеалогическое древо Султанов Османской империи: Османские правители, вначале, будучи еще бейлербеями Анатолии, женились на дочерях византийских императоров...
Интересное:
Финансовый рынок и его значение в управлении денежными потоками на современном этапе: любому предприятию для расширения производства и увеличения прибыли нужны...
Аура как энергетическое поле: многослойную ауру человека можно представить себе подобным...
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Дисциплины:
|
из
5.00
|
Заказать работу |
Вернемся еще раз к явлению трансформации, с которой мы уже познакомились в предыдущей главе. Мы видели, что колонии пневмококков подвержены диссоциации, которая проявляется в изменении их свойств. Клетки пневмококков формы S очень вирулентны (вызывают типичную форму воспаления легких). Они образуют слизистые капсулы, которые обычно прикрывают пары клеток. Эти капсулы состоят из сложного полисахарида. Напротив, пневмококки формы R, образовавшиеся в результате диссоциации формы S, не вирулентны, не образуют капсул и не располагаются парами. Эти различия проявляются во время опытов на животных (определение вирулентности), микроскопирования (по присутствию парнорасположенных клеток с капсулами) и культивирования на питательной среде – агаре (по форме и цвету колоний).
Пневмококки доставили ранее много беспокойств медикам, поскольку врачи еще не располагали пенициллином и другими современными средствами борьбы с воспалением легких. В 1928 году английский бактериолог Ф. Гриффит открыл явление трансформации у этих микроорганизмов. Гриффит испытывал влияние невирулентных R‑пневмококков на подопытных мышей. Он привил им миллионы клеток типа R, и мыши погибли. Исследуя органы погибших мышей в поисках этих бактерий, ученый столкнулся с неожиданным фактом: вместо лишенных капсулы пневмококков типа R там были исключительно имевшие капсулы вирулентные пневмококки типа S.
Гриффит предположил, что часть клеток, погибая, выделяет какое‑то вещество, которое заставляет оставшиеся в живых клетки R образовывать капсулы. Это предположение он решил проверить следующими опытами. В организм мыши вносилось небольшое количество R‑клеток вместе с многочисленными, но умерщвленными высокой температурой S‑клетками. Клетки S принадлежали к так называемому типу I. Позднее, как и ожидал Гриффит, из мышей были выделены живые S‑клетки. Во втором опыте он использовал живые R‑клетки типа I и умерщвленные S‑клетки типа II. В этом случае он получил из мышей живые S‑клетки типа II. Напрашивалось единственно возможное объяснение: нечто из умерщвленных S‑клеток типа II превратило (трансформировало) их родственников – лишенные капсулы R‑клетки типа I – в вирулентные S‑клетки, способные образовывать капсулы типа II. Чем же было это «нечто», что вызвало трансформацию? Гриффит назвал его трансформирующим началом.
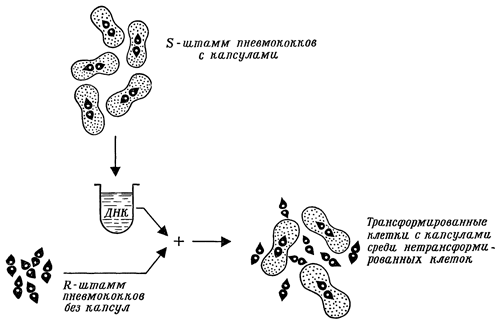
Схема трансформации пневмококков R‑формы в S‑форму при помощи ДНК, выделенной из клеток S и перенесенной к клеткам R
Это «нечто», обладающее способностью активного действия, привлекло внимание исследователей, и не удивительно, что К. Т. Эйвери со своими коллегами из Рокфеллеровского университета попытался вскрыть его сущность. В 1944 году ученые с достоверностью доказали, что трансформирующим началом была ДНК из S‑клеток. Читателю, уже немало узнавшему о генетической роли ДНК, такой результат представляется довольно очевидным. Но те, кто впервые с ним столкнулся, были в менее выгодном положении.
Первой задачей исследователей было вырастить достаточное количество клеток вирулентных пневмококков, чтобы получить из них трансформирующее начало. После их умерщвления и разрушения при помощи дезоксихолата натрия (вещества, содержащегося в желчи) они получили около 1 мг чистой ДНК из 30 миллиардов клеток. Если воздействовать этим веществом на R‑клетки, то в их потомстве 1 % клеток начинает образовывать капсулы, и это свойство наследственно передается из поколения в поколение. Трансформация, таким образом, вызывалась при помощи ДНК из вирулентных пневмококков. Это позволило исследователям сделать вывод, что ДНК является трансформирующим началом.
Последовали подтверждения из многих микробиологических лабораторий мира, где также удалось провести трансформацию бактерий, но уже других, в частности кишечной палочки Escherichia coli. Трансформирующим началом во всех случаях была ДНК.
Однако, пневмококки в этом отношении занимали, как оказалось, особое положение, поскольку у них удалось трансформировать более 25 наследственных признаков, среди которых была способность к образованию капсул из полисахаридов, а также к созданию особых типов колоний и устойчивость к лекарственным препаратам. При трансформации обычно переносится лишь один из наследуемых признаков даже в том случае, если используется ДНК из пневмококков, обладающих двумя или тремя отличительными признаками. Если, например, на 1 000 000 клеток пневмококка формы R действовать десятимиллионной частью грамма ДНК из пневмококка формы S, устойчивой к пенициллину и стрептомицину, то трансформируется лишь 50 000 клеток. Из них приблизительно 49 000 получат одно из переносимых свойств (способность образовывать капсулы или устойчивость к определенному лекарству), около 800 клеток приобретут два новых свойства и только 4 клетки получат все три новых признака: способность к образованию капсул, устойчивость к пенициллину и устойчивость к стрептомицину.
ДНК при трансформации нередко передает и такие свойства, которые долгое время могут оставаться скрытыми и проявляются лишь при особых обстоятельствах. Так, например, трансформацией объясняется передача способности к созданию фермента, вызывающего окисление маннита (вещества, близкого к сахару маннозе). Этот фермент проявится только в том случае, если мы будем выращивать пневмококки на питательной среде, где единственным источником углерода будет маннит. Трансформированные пневмококки начнут окислять маннит и использовать его как источник углерода и энергии, тогда как их нетрансформированная «родня» в этой среде не будет расти и размножаться.
Интересно, что однажды трансформированные бактерии не теряют в дальнейшем способности к новой трансформации, особенно к такой, которая совершается с помощью ДНК, обладающей многими закодированными свойствами.
Читатель познакомится также и с такими важными проблемами, какой является, например, в наше время – в эру антибиотиков – устойчивость некоторых болезнетворных микробов к антибиотикам. Если, например, стафилококки стали устойчивыми к пенициллину, то лечение им инфекционных заболеваний, вызванных этими стафилококками, окажется бесполезным. Американский естественнонаучный журнал Science опубликовал в 1962 году сообщение о том, что ДНК, содержащаяся в восприимчивых к пенициллину стафилококках, может трансформировать устойчивые к нему стафилококки в чувствительные. К сожалению, такую трансформацию трудно осуществить в большом масштабе, иначе мы смогли бы трансформировать устойчивых представителей стафилококков в чувствительные и затем уничтожить их пенициллином!
Бактериофаги в действии
Мы уже познакомились со строением вирусов. Узнали мы и об их сходстве с неживыми химическими соединениями. Теперь нам предстоит подробнее рассмотреть особенности, связывающие их с живыми организмами.
Итак, мы уже знаем, что вирусы сходны с последними прежде всего способностью размножаться. Однако их размножение происходит лишь в живых клетках хозяина. Сведения о том, как вирус проникает в клетку и как производит в ней разрушительные изменения, впервые были получены при наблюдениях над бактериофагами – врагами кишечной бактерии Escherichia coli. Электронный микроскоп помог нам раскрыть тайну внешнего строения бактериофага Т4, изображенного на фото 49. Ниже дана схема строения фага Т2. Частица фага имеет головку, напоминающую шестигранную призму с шестигранными же крышечками с обеих сторон. От головки отходит продолговатый хвостовой придаток, конец которого снабжен несколькими нитевидными «щупальцами». В придатке имеется канал, который соединяет его с головкой и в котором находится макромолекула ДНК. Остальные части оболочки фага – белковой природы.
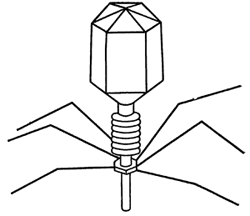
Модель бактериофага Т2 – паразита‑бактерии Escherichia coli. Граненая головка соединена с хвостовым придатком, имеющим внутренний канал и выросты для прикрепления к бактерии. На модели хвостовой придаток сжался и из него выдвинулась трубка с каналом, по которому ДНК проникает в бактерию.
Бактериофаг Т2 не имеет, по существу, двигательных органов и в жидкой среде перемещается пассивно, в результате столкновения с молекулами среды. Таким образом, он приходит в соприкосновение и с бактерией. Фаг «пристает» к ее поверхности при помощи щупалец. В конце хвостового придатка содержится фермент, поражающий оболочку бактерии и «проедающий» в ней небольшое отверстие. После этой операции придаток втягивается (укорачиваясь подобно гармонике) в тело фага.
ДНК из головки бактериофага через канал в хвостовом придатке и через образованное отверстие в стенке бактерии проникает в клетку. Все это несколько напоминает процедуру инъекции при помощи шприца. Белковая оболочка головки и придатка остаются снаружи, а в клетку бактерии входит только ДНК. И начинается драматическая фаза – «саморазмножение» ДНК фага. Макромолекула ДНК «принуждает» бактерию участвовать в процессе своего размножения, используя ее «сырье» и весь ферментный аппарат.
Как мы уже знаем, молекула ДНК образует подобие двойной спирали, состоящей из двух свернутых цепочек. При ее размножении эти цепочки, развертываясь, освобождаются друг от друга и к каждой из них присоединяются из среды основные структурные единицы (нуклеотиды), дополняя недостающую половину молекулы. Из первичной макромолекулы ДНК возникают две дочерние, а со временем их раскрученные цепочки дополняются новыми нуклеотидами. В результате появляются уже 4 макромолекулы, и вскоре число их возрастает до 8, 16 и т. д. По прошествии получаса из одной первичной молекулы ДНК фага в клетке находятся уже 150–300 ее потомков, принуждающих бактерию синтезировать белок для головки и хвостового придатка бактериофага. Клетка лопается, как воздушный шарик, выпуская в среду 150–300 бактериофагов, и те стремительно нападают на следующие бактерии. Процесс повторяется, и еще через полчаса в среде уже десятки тысяч фагов. Спустя некоторое время там не остается ни одной живой бактерии, кишат лишь победившие бактериофаги.
Весь этот процесс уничтожения бактерий и размножения фагов можно наблюдать под электронным микроскопом (фото 50). Впрочем, результаты такого уничтожения можно увидеть и невооруженным глазом. С этой целью в чашку Петри с агаризованной питательной средой наносят несколько капель культуральной жидкости, содержащей бактерии. Внесем туда же несколько капелек из другой жидкой среды, в которой находятся бактериофаги. Перемешаем их и равномерно нанесем несколько капель этой смеси на поверхность агара. Затем чашку Петри поместим в термостат при температуре 37 °C. Через 24 часа мы обнаружим следующую картину.
Несколько сотен бактериофагов, помещенных на поверхность агара, уже вошли в контакт с клетками бактерий и сделали свое дело. Там же находятся и клетки, избежавшие нападения фагов и продолжающие нормально размножаться. На поверхности питательной среды образуется видимый невооруженным глазом налет, местами прерванный округлыми пятнами правильной формы, на которых бактерии отсутствуют. Эти пятна – свидетельство уничтожающей деятельности бактериофагов (фото 51).

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Своеобразие русской архитектуры: Основной материал – дерево – быстрота постройки, но недолговечность и необходимость деления...

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!