Vasiliy A. Kolbin
Vishersky nature reserve
36-B Gagarin Str, Krasnovishersk Perm region Russia 618590
e-mail: [email protected]
Daria V. Kolbina
Vishersky nature reserve
36-B Gagarin Str, Krasnovishersk Perm region Russia 618590
e-mail: [email protected]
THE BIRD COMMUNITIES IN PLACES OF TRADITIONAL
NATURE USE MUNCIE
Аннотация. Описаны сообщества птиц подгольцового и горно-тундрового пояса заповедника «Вишерский», которые в той или иной степени подвергались воздействию оленеводства. Даны количественные и качественные характеристики орнитокомплексов.
Ключевые слова: сообщества птиц, заповедник «Вишерский», фауна, численность видов.
Abstract. Bird communities of the subalpine belt and mountain-tundra belt of the Vishersky reserve, which were affected by reindeer husbandry to varying degrees, are described. Quantitative and qualitative characteristics of ornithocomplexes are given.
Key words: bird communities, Vishersky reserve, fauna, number of species.
Изучение любого вида в природе едва ли возможно без понимания его взаимоотношений с другими видами. Взаимодействующие между собой виды (популяции) определенной территории – в нашем случае определенного фитоценоза – образуют сообщество. Обычно подразумевается, что организмы, слагающие сообщество, принадлежат к одному трофическому уровню [4]. Но выделенные и описанные геоботаниками сообщества растений (фитоценозы) часто являются слишком мелкими специфическими пространственными единицами, среди которых такие высокоподвижные животные, как птицы, перемещаются совершенно свободно. В природе нередко приходится обнаруживать вроде бы чисто дендрофильных птиц, например поползней или кедровок, в горной тундре – среди останцев, поэтому орнитологу приходится оперировать крупными сообществами, возможно, делая слишком грубые построения с точки зрения фитоценологии. Невозможность вычленения соответствующей группировки птиц для каждой формации, выделяемой геоботаниками, отмечал также А.А. Назаренко [9].
В настоящее время большинством исследователей признается, что сообщества чаще всего плавно сменяют друг друга в соответствии с градиентом условий. Поэтому правильнее говорить о континууме сообществ, а не об их дискретных типах. Но на практике классификация сообществ оказывается необходимой для решения множества задач: например, выявления взаимосвязей между различными совокупностями локальных популяций видов, направленности их эволюции, поиска сходства и различия между ними и многих других. При этом понятно, что каждое выделенное нами сообщество – это удобная абстракция, а не конкретная реальность. Уместно вспомнить слова Р. Уиттекера [14]: «Никто еще не утверждал, что нельзя пользоваться названиями оттенков цвета только потому, что они являются субъективно различаемыми частями непрерывного цветового спектра».
Характеристика сообщества только через создание перечня видов, формирующих его, является предельно упрощенной. Более законченная картина получается при учете значимости вида (его ранга) в сообществе. Для этого целесообразно построение диаграмм рангового распределения видов по обилию, биомассе или другим показателям [2].
В первой половине XX в. анализ распространения видов в пространстве позволил зоогеографам сформулировать концепцию исторически сложившихся комплексов животного мира (типов фауны), которые объединяются общностью области распространения [3]. В дальнейшем при отнесении того или иного вида к определенной фауне учитывались экологические связи с растительностью, широтная, меридиональная и региональная зональности распределения организмов [6, 7]. В конце XX в. в экологии произошла смена парадигм: организмистскую – куда прекрасно вписывались представление о существовании дискретных фаун – сменила континуальная [8]. Но несостоятельность концепции фаун никем не была доказана, а привлекательность ее основополагающих принципов не исчезла и в настоящее время. Поэтому при изучении сообществ животных по-прежнему целесообразно наряду с другими аспектами рассматривать их и с позиции экологической зоогеографии.
С другой стороны, нельзя забывать, что современное состояние распределения организмов в регионе и отдельном сообществе – это временной срез или, по словам Б.К. Штегмана [15], «моментальный снимок» непрерывного процесса его изменения.
Б.К. Штегманом [16,17] для Евразии выделен ряд типов фаун, из которых на территории ООПТ обнаружены представители пяти комплексов: европейского, сибирского, китайского, арктического и транспалеарктов.
Целью данной работы являлось изучение сообществ птиц, сформировавшихся в местах традиционного природопользования манси в подгольцовом и горно-тундровом поясах на территории Вишерского заповедника (рис. 1). В настоящее время, когда уклад манси изменился, и домашние олени исчезли, собранные данные представляют собой исторический интерес. Кроме того, они могут использоваться как сравнительный материал для дальнейших исследований.
Материалы и методы. Сбор данных по населению птиц осуществлялся на территории Вишерского заповедника. Полевые исследования проводились в летний период 1994–1996, 1999, 2001–2002, 2004–2016 гг. При этом записывались все птицы, обнаруженные визуально или на слух.
Маршрутные учеты проводились с использованием наиболее распространенных методик [10, 18]. Во время пеших маршрутов учитывались все встреченные птицы, для каждого вида на основе нескольких промеров определялась максимальная дальность обнаружения и рассчитывалась полоса учета. По этим данным вычислялась плотность населения вида. В гнездовое время она оценивалась в парах на км2. При этом нетерриториальные виды объединялись в пары условно.
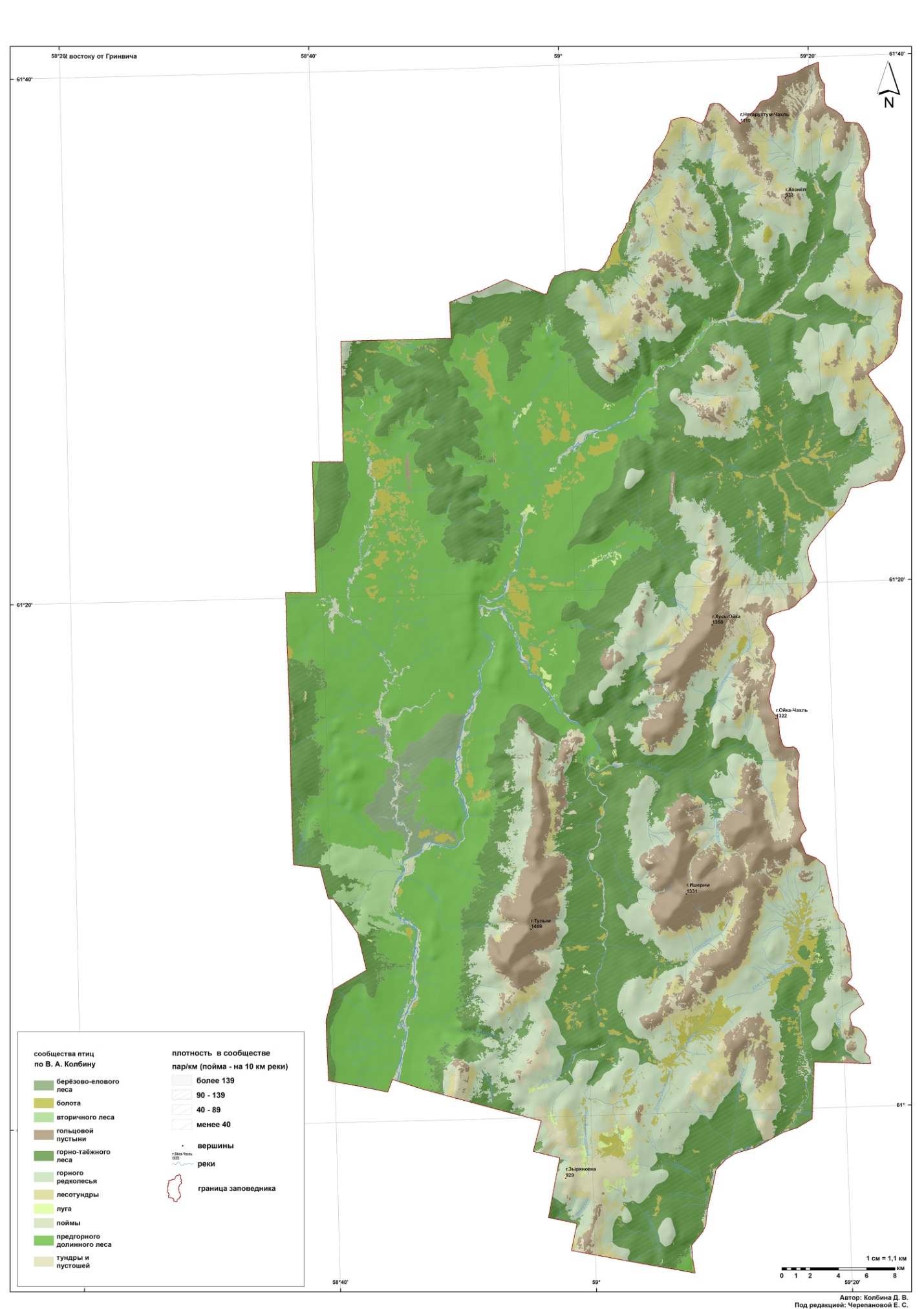
Рис. 1. Размещение сообществ птиц на территории заповедника «Вишерский»
Для тех видов, которые многократно учитывались в разные годы, рассчитывалась средняя плотность населения, для оценки варьирования данных вычислялось стандартное отклонение. По результатам учетов строились диаграммы рангового распределения птиц в сообществах по обилию. Для характеристики сообществ рассчитывались индексы полидоминантности [12], или индексы разнообразия Симпсона [2]:
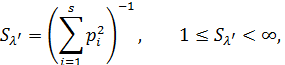
где pi – доля i -го вида в генеральной совокупности, включающей S видов, или вероятность его встречи в соответствующем населении.
Выровненность сообщества оценивалась по формуле
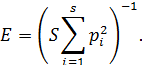
Степень сходства различных сообществ оценивалась по индексу Чекановского – Съеренсена [12]. Для качественных данных:
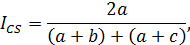
где a – число видов, входящих в оба списка, b и c – число видов, входящих только в 1-й или во 2-й списки.
Для количественных данных:
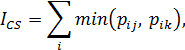
где pij – доля i -го вида в выборке J, а pik – доля i -го вида в выборке K. Полученные матрицы сходства были преобразованы в матрицы дистанций (как 1 – Ics), которые обрабатывались методами кластерного анализа.
Результаты и их обсуждение. Обширные площади в подгольцовом поясе занимают горные пустоши, с доминированием лерхенфельдии или щучки горной Avenella flexuosa, овсяницы рупрехта Festuca ruprechtii и осоки мечелистной Carex ensifolia. Понятие «горных пустошей» для Вишерского Урала введено А.М. Овесновым [11], где этот тип растительности является вторичным. Он сформировался на месте мохово-лишайниковых тундр под воздействием обогащения почв продуктами жизнедеятельности оленей и человека. Подгольцовые заросли кустарников представлены вересовниками – зарослями столообразных кустов можжевельника сибирского Juniperus sibirica, ивняками –зарослями ив Salix glauca, S. Lapponum, S. lanata, ерниками – зарослями карликовой березки Betula nana. Вдоль верхней границы леса часто располагаются обширные осоково-сфагновые болота с мочажинами, чередующиеся с языками березового криволесья. Такие болота особенно заметны в верховьях р. Малая Мойва вдоль склона хр. Муравьиный Камень. Растительность горно-тундрового пояса представлена широким спектром тундровых формаций: это лишайниковые, моховые, травяные, кустарничковые и смешанные кустарничково-лишайниковые, кустарничково-моховые, кустарничково-травяно-моховые тундры.
Население птиц открытых ландшафтов подгольцового и горно-тундрового пояса достаточно резко отличается от лесных формаций, что отражает значительное изменение состава и структуры фитоценозов. С набором высоты происходит упрощение и обеднение сообществ, но при этом видовой состав их становится наиболее своеобразным.
Нелесные горные орнитокомплексы объединяет количественное доминирование одного вида, поэтому их можно условно обозначить как «сообщества лугового конька».
Сообщество лесотундры. Очевидно, что наибольшее видовое разнообразие и плотность населения птиц наблюдается в переходных фитоценозах, которые прилегают к редколесьям и криволесьям, и обозначены нами как сообщество лесотундры или кустарниковой тундры у других авторов [5]. Оно охватывает вересовники, ерники и ивняки и располагается в диапазоне абсолютных высот от 700–750 до 800 м. На пологих склонах площадь таких кустарниковых зарослей бывает достаточно значительной. Для них характерна высокая мозаичность растительности [1, 13]. Заросли можжевельника здесь соседствуют с куртинами карликовой березы, в увлажненных местах их пронизывают ивняки, кое-где еще произрастают одиночные деревья или группы их. К вересовникам приурочены щучковые пустоши, которые среди кустарников выглядят как небольшие или обширные поляны. С подъемом в горы высота кустарников уменьшается.

Рис. 2. Распределение птиц по обилию в сообществе птиц лесотундры
В данном сообществе выявлено 22 вида птиц, у 12 из них доля участия превысила 1 % (рис. 2). Суммарная средняя плотность населения здесь составила 118 пар/км2. Безоговорочно доминирует луговой конек Anthus pratensis – 46,2 %. О других массовых видах можно говорить только как о субдоминантах: в этой группе стабильно присутствуют пеночка-весничка – 14,1 %, варакушка – 11,5 %, овсянка-крошка – 6,7 %. Куриные птицы представлены тетеревом Lyrurus tetrix – 0,9 % и белой куропаткой Lagopus lagopus – 0,7 %. Несколько особняком стоит население птиц высоких – до 2,5 м – ивняков, которые обычно занимают ложа ручьев и достаточно глубоко вклиниваются в горные пустоши и тундры. Именно в ивняках или рядом с ними обитают в подгольцовом и горно-тундровом поясах садовая камышевка Acrocephalus dumetorum, северная бормотушка Hippolais caligat a, серая Sylvia communis и садовая славки S. borin, славка-мельничек S. curruca. Впрочем, этот комплекс видов связан с данной формацией также в предгорьях и на равнине. С понижением высоты ивняков до 0,5–1,0 м население птиц утрачивает своеобразие и становится типичным сообществом лугового конька.
Индекс полидоминантности составил здесь 3,9, выровненность – 0,18. Данные показатели подчеркивают разительные отличия тундровых сообществ от лесных орнитокомплексов (табл. 1). Также это подтверждают результаты кластерного анализа (рис. 5–6).
Осоково-сфагновые болота. Другим орнитокомплексом лугового конька, соседствующим с криволесьями и редколесьями, является сообщество птиц осоково-сфагновых болот. Такие формации обследовались нами в верховьях р. Малая Мойва вдоль юго-восточного склона хр. Муравьиный Камень, на водоразделе рек Большая Мойва и Велс, а также в пойме р. Большая Мойва. Здесь также выявлено 22 вида, у 13 доля участия превысила 1% (рис. 3). Суммарная средняя плотность населения составила 90 пар/км2. Доминирование лугового конька – 23,4% в этом сообществе не столь ярко выражено, как в кустарниковых формациях, доля чечетки не многим меньше – 21,6%, но, как уже отмечалось, численность чечетки подвержена значительным колебаниям. Только в этой формации регулярно регистрировались желтые Motacilla flava – 6,3% и желтоголовые M. citreola – 1,4% трясогузки. Следует отметить, что кулики – фифи Tringa glareola и большой улит T. nebularia – встречались только на болотах вблизи рек или озер.

Рис. 3. Распределение птиц по обилию в сообществе птиц осоково-сфагновых болот
По показателям разнообразия данный орнитокомплекс занимает промежуточное положение между лесными и тундровыми сообществами (табл. 1). По индексу Чекановского – Съеренсена наблюдается наибольшее сходство с сообществами птиц лесотундры и редколесий (табл. 2).
Сообщество тундры и пустошей. Население птиц разнообразных растительных формаций открытых горных ландшафтов с кустарничками, низкими кустарниками или без таковых достаточно однотипно, оно обозначено нами, как сообщество тундры и пустошей. Сообщество охватывает лишайниковые, моховые, травяные, кустарничковые и смешанные кустарничково-лишайниковые, кустарничково-моховые, кустарничково-травяно-моховые тундры, а также щучковые, овсяницевые и осоковые пустоши. В орнитокомплексе насчитывается 14 видов, у 9 из которых доля участия превысила 1 % (рис. 4). Суммарная средняя плотность населения – 40 пар/км2. Доминирование лугового конька проявляется в этом комплексе наиболее сильно – 61,5 %. Возрастает значимость птиц, не относящихся к отряду воробьинообразных: белая куропатка – 8,6 %, тундряная куропатка Lagopus mutus – 6,1 %, золотистая ржанка Pluvialis apricaria – 5,3 %, бекас Gallinago gallinago – 2,5 %. Весьма заметными становятся вóроны Corvus corax – 0,9 %. Хищники: дербник Falco columbarius – 0,5 %, пустельга F. tinnunculus – 0,4 %, полевой лунь Circus cyaneus – 0,4 % – охотятся во всех сообществах открытых ландшафтов, а гнезда устраивают на останцах или среди курумов, поэтому их «приписывание» к сообществу тундры достаточно условно.

Рис. 4. Распределение птиц по обилию в сообществе птиц тундры и пустошей
Как и можно было предполагать, показатели разнообразия в этом сообществе имеют самые низкие значения (см. табл. 1). Сходство по индексу Чекановского – Съеренсена наиболее высоко с близкими сообществами лесотундры и болот и ничтожно – с лесными (табл. 2).
Самым «высоким» и самым «простым» орнитокомплексом является сообщество каменистой тундры, которое можно рассматривать и как часть сообщества тундры, поскольку в нем нет новых видов. Каменистая тундра распространена на основных хребтах, она проникает в гольцовые пустыни и соседствует с ними. Основные учеты в данной формации проводились на хр. Ольховочный, где каменистые тундры занимают плоские вершины и плато на севере, а также на хр. Молебный и Ишерим. Сообщество насчитывает четыре вида: луговой конек – 63,5 %, тундряная куропатка – 15,3 %, каменка Oenanthe oenanthe – 10,9 %, золотистая ржанка – 10,2 %. Средняя суммарная плотность – 18 пар/км2. Сообщество имеет сходство с тундровыми сообществами птиц и не имеет с лесными (см. табл. 2).
Рассмотренные орнитокомплексы явно делятся на две группы: группу лесных сообществ и группу сообществ открытых ландшафтов. Это подтверждают показатели разнообразия (см. табл. 1), индексы сходства (см. табл. 2) и кластерный анализ (рис. 5–6).
С ростом высоты над уровнем моря происходит снижение значений индекса полидоминантности (см. табл. 1), причем, если в пределах лесных сообществ изменения не значительны, то при переходе от редколесий к болотам и лесотундре виден резкий скачок. Изменения показателей выровненности не столь однозначны, но в целом также укладываются в схему снижения разнообразия и упрощения орнитокомплексов с набором высоты. Самый высокий показатель выровненности в каменистой тундре объясняется малым количеством видов – их всего четыре, поэтому им можно пренебречь, тем более, что данный комплекс можно рассматривать как часть сообщества тундры.
Сообщества птиц открытых горных ландшафтов, обозначенные нами как сообщества лугового конька, образуют вторую ветвь на дендрограмме (см. рис. 5–6). Они характеризуются при сравнении с лесными формациями упрощенностью, доминированием одного вида и высоким своеобразием. Последний факт наглядно демонстрирует полное отсутствие перекрывания с лесными формациями по индексу Чекановского – Съеренсена сообщества каменистой тундры и ничтожное перекрывание сообщества тундры и пустошей (см. табл. 2). Только здесь обитают такие виды, как золотистая ржанка, белая и тундряная куропатки, хрустан Eudromias morinellus, лапландский подорожник Calcarius lapponicus. В самом «верхнем» сообществе – сообществе каменистой тундры наблюдаются самые низкие значения плотности населения – 18 пар/км2.
Таблица 1
Показатели разнообразия сообществ птиц Вишерского заповедника
| Сообщество
| Индекс полидоминантности
| Выровненность
|
| Долинные леса
| 20,51
| 0,48
|
| Горно-таежные леса
| 20,71
| 0,41
|
| Редколесья
| 16,67
| 0,34
|
| Пойма, лес и акватория
| 19,32
| 0,20
|
| Пойменные луга
| 11,01
| 0,52
|
| Осоково-сфагновые болота
| 7,36
| 0,33
|
| Лесотундра
| 3,90
| 0,18
|
| Тундра и пустоши
| 2,50
| 0,18
|
| Каменистая тундра
| 2,23
| 0,56
|
Таблица 2
Перекрывание сообществ птиц по индексу Чекановского – Съеренсена (I cs)
| Тундра и пустоши
| Лесотундра
| Осоково-сфагновые болота
| Луга
| Редколесья
| Горно-таежные леса
| Долинные леса
| Пойма
|
| Каменистая тундра
| 0,760
| 0,478
| 0,240
| 0
| 0
| 0
| 0
| 0
|
| Тундра и пустоши
| 0,498
| 0,341
| 0,068
| 0,051
| 0,062
| 0,045
| 0,042
|
| | Лесотундра
| 0,514
| 0,036
| 0,325
| 0,150
| 0,136
| 0,133
|
| | | Осоково-сфагновые болота
| 0,137
| 0,334
| 0,118
| 0,109
| 0,129
|
| | | | Луга
| 0,072
| 0,057
| 0,149
| 0,179
|
| | | | | Редколесья
| 0,710
| 0,586
| 0,419
|
| | | | | | Горно-таежные леса
| 0,707
| 0,426
|
| | | | | | | Долинные леса
| 0,566
|

Рис. 5. Дендрограмма сходства сообществ птиц по обилию на основе дистанций Ics

Рис. 6. Дендрограмма сходства сообществ птиц по видовому составу на основе дистанций Ics
Своеобразие сообщества птиц тундры и пустошей (как центрального среди орнитокомплексов горных открытых ландшафтов) проявляется и в фаунистическом аспекте. Так, здесь наблюдается доминирование представителей европейской фауны по обилию – 61 % (рис. 7), поскольку главный доминант – луговой конек отнесен Б.К. Штегманом к европейской фауне. По количеству видов в сообществе преобладают транспалеаркты – 57 %, а на второй позиции оказываются виды арктического комплекса – 29 % (рис. 8).

Рис. 7. Соотношение (%) фаунистических комплексов по обилию представителей в сообществе птиц тундры и пустошей

Рис. 8. Соотношение (%) фаунистических комплексов по количеству видов в сообществе птиц тундры и пустошей
Список литературы
1. Белковская Т.П., Безгодов А.Г., Овеснов С.А. Сосудистые растения Вишерского заповедника. – Пермь, 2004. – С. 1–103.
2. Бигон М., Харпер Дж., Таунсенд К. Экология. Особи, популяции и сообщества. – М., 1989. –Т. 2. – С. 1–477.
3. Бобринский Н.А., Зенкевич Л.А., Бирштейн Я.А. География животных. – М., 1946. –256 с.
4. Гиляров А.М. В поисках универсальных закономерностей организации сообществ: прогресс на пути нейтрализма // Журнал общей биологии. – 2010. – Т. 71, № 5. – С. 386–401.
5. Головатин М.Г., Пасхальный С.П. Птицы Полярного Урала. – Екатеринбург, 2005. – С. 1–560.
6. Куренцов А.И. Зоогеография Приамурья. – М.–Л. 1965. –154 с.
7. Матюшкин Е.Н. Региональная дифференциация лесной фауны Палеарктики в прошлом и настоящем // Теоретические и прикладные аспекты биогеографии. –М.: Наука, 1982. – С. 59–80.
8. Миркин В.М. О парадигмах в фитоценологии // Журн. общ. биол. – 1984. – Т. 45, № 6. – С. 749–758.
9. Назаренко А.А. Птичье население смешанных и темнохвойных лесов Южного Приморья, 1962– 1971 гг. // Фаунистика и биология птиц юга Дальнего Востока. Владивосток, 1984. – С. 60–70.
10. Наумов Р.П. Методики абсолютного учета птиц в гнездовой период на маршрутах // Зоологический журнал. – 1965. – №1. – С. 81–92.
11. Овеснов А.М. Горные луга Вишерского Урала // Тр. ЕНИ при Перм. ун-те. – 1948. –Т. 10, вып. 1. – С. 1–86.
12. Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. – М., 1982. – С. 1–287.
13. Растительность и флора, грибы, лишайники заповедника «Вишерский» / Белковская Т.П. [и др.]. – Соликамск, 2014. – С. 1–400.
14. Уиттекер Р. Сообщества и экосистемы. – М., 1980. – С. 1–327.
15. Штегман Б.К. О путях формирования фаун и методике их изучения // Изв. Всесоюз. геогр. о-ва. – 1950. – Т. 82, № 4. – С. 392–395.
16. Штегман Б.К. Основы орнитогеографического деления Палеарктики // Изв. АН СССР. Серия: Биология. – 1936. – № 2–3. – С. 523–563.
17. Штегман Б.К. Основы орнитографического деления Палеарктики. – М. – Л.: Изд-во АН СССР, 1938. – 156 с.
18. Щеголев В.И. Количественный учет птиц в лесной зоне // Методики исследования продуктивности и структуры птиц в пределах их ареалов. – Вильнюс, 1977. – С. 95–102.
УДК 598.2(470.53)
DOI 10.24411/2308-720Х-2019-10004
Колбина Дарья Васильевна, Колбин Василий Анфимович ·
ФГБУ «Государственный заповедник “Вишерский”»,
Пермский край, 618590, г. Красновишерск, ул. Гагарина, 36 б
e - mail: dashakolbina 28@ gmail. com; kgularis @ mail. ru





