Список сокращений
Введение
Обзор литературысинтетаза и атерогенез
Общая характеристика гена eNOS
Материалы и методы
Обследуемые группы
Использованные материалы, реактивы и оборудование
Выделение ДНК из лейкоцитов периферической крови
Полимеразно - цепная реакция (ПЦР)
Проведение ПЦР для 4a4b полиморфизма гена эндотелиальной NO-синтазы (eNOS)
Электрофорез
Статистический анализ
Результаты и обсуждение
Выводы
СПИСОК ЛИТЕРАТУРЫ
Список сокращений
Русскоязычные
ССП - сердечно-сосудистая патология
ИБС - ишемическая болезнь сердца
АГ - артериальная гипертензия
ИМ - инфаркт миокарда
ДНК - дезоксирибонуклеиновая кислота
п.н. - пар нуклеотидов
п.о. - пар оснований
ПЦР - полимеразная цепная реакция
СД2 - сахарный диабет тип 2
ЭДТА - этилендиаминтетрауксусная кислота
ЛПНП - липопротеиды низкой плотности
ЛПВП - липопротеиды высокой плотностиЗ - сердечно-сосудистые заболевания
ОР - относительный риск
ДИ - доверительный интервал
Англоязычные
- оксид азота - эндотелиальная NO-синтетаза
Введение
Эндотелиальные клетки первыми встречаются с реактивными свободными радикалами, с окисленными липопротеинами низкой плотности, с гиперхолестеринемией, с высоким гидростатическим давлением внутри выстилаемых ими сосудов (при артериальной гипертонии), с гипергликемией (при сахарном диабете). Все эти факторы приводят к повреждению эндотелия сосудов, к дисфункции эндотелия, как эндокринного органа и к ускоренному развитию ССЗ.
Из всех факторов, синтезируемых эндотелием, роль "модератора" основных функций эндотелия принадлежит эндотелиальному фактору релаксации или оксиду азота (N0). Именно это соединение регулирует активность и последовательность "запуска" всех остальных биологически-активных веществ, продуцируемых эндотелием. Оксид азота не только вызывает расширение сосудов, но и блокирует пролиферацию гладкомышечных клеток, препятствует адгезии клеток крови и обладает антиагрегантными свойствами. Таким образом, оксид азота является базовым фактором антиатерогенеза[1].
Ген эндотелиальной NO-синтетазы (eNOS), отвечает за синтез оксида азота (NO) эндотелием и является ключевым ферментом в регуляции тонуса кровеносных сосудов, в работе гладкомышечной мускулатуры сосудистой стенки и процессов тромбообразования. Полиморфизм в гене eNOS может стать прогностически важным в формировании группы риска сердечно-сосудистой патологии (ССП) среди детей с отягощённой наследственностью по ССЗ. Поэтому целью нашего исследования стало определение роли полиморфизма eNOS в формировании риска ССП у детей с отягощённым анамнезом по ССЗ.
Для осуществления нашей цели были поставлены следующие задачи:
Провести анализ наследственной отягощенности у детей в группе риска по ожирению, по сахарному диабету тип 2 (СД2) и по сердечно-сосудистой патологии.
Генотипировать банк ДНК всех детей по полиморфизму 4a/4b гена eNOS.
Сравнить частоты встречаемости аллелей и генотипов по данному гену между группами и в зависимости от пола и типа отягощенной наследственности, а так же в зависимости от количества у исследованных детей близких родственников с ССЗ, ожирением и наличием СД2.
Работа проводилась в лаборатории молекулярной диагностики с расширенной группой по экогенетике СПбГПМА под руководством д.м.н., проф., В.И. Ларионовой.
Обзор литературы
NO-синтетаза и атерогенез
Основным триггерным фактором при атеросклерозе считается окисление липопротеидов низкой плотности (ЛПНП) свободными радикалами. Окисленные липиды, входящие в состав ЛПНП, активируют макрофаги. Макрофаги начинают продуцировать цитокины и свободные радикалы. Свободные радикалы окисляют NO до нитратов, а цитокины индуцируют экспрессию и синтез индуцибельной NO-синтетазы в гладкомышечных эндотелиальных клетках. Индуцибельная NO-синтетаза создает относительный дефицит субстрата, или аргинина, поскольку она оттягивает его как бы на себя и уменьшает его доступность для эндотелиальной NO-синтетазы [2]. Известно, что у больных атеросклерозом уровень аргинина в плазме крови снижен приблизительно на 30 %.
В настоящее время известно, что эндотелий регулирует сосудистый тонус через освобождение сосудорасширяющих и сосудосуживающих факторов и модулирует сократительную активность гладкомышечных клеток. К эндотелиальным факторам дилатации относится и оксид азота. NO является основным вазодилататором, препятствующим тоническому сокращению сосудов нейронального, эндокринного или локального происхождения.
В физиологических условиях NO постоянно вовлечен в адаптацию сосудистой системы к повышенным метаболическим потребностям, физическим нагрузкам. Избыток NO отвечает за увеличение периферической вазодилатации при вазоплегическом шоке, а недостаток NO может приводить к тяжелым заболеваниям, включая артериальную гипертонию, ишемическую болезнь сердца и атеросклероз [2]. синтетаза (NOS), принимая участие в синтезе оксида азота (NO) эндотелием, является ключевым ферментом в регуляции тонуса кровеносных сосудов, в работе гладкомышечной мускулатуры сосудистой стенки и процессов тромбообразования [3]. NO-синтетаза существует в виде трех основных изоформ, которые получили свое название по типу клеток, в которых они были впервые обнаружены: нейрональная NO-синтетаза (nNOS или NOS I), эндотелиальная NO-синтетаза (eNOS или NOS III) и NO-синтетаза макрофагов или индуцибельная NO-синтетаза (iNOS или NOSII). Нейрональная и эндотелиальная NO синтетазы являются ферментами со стабильной активностью, в то время как активность макрофагальной или индуцибельной NO- синтетазы в большей степени регулируется цитокинами. Эндотелиальная NO- синтетаза стабильно экспрессируется в эндотелиальных клетках [4].
Ингибирование NO-синтетазы приводит ко всем органическим последствиям тяжелой и продолжительной артериальной гипертензии, включая атеросклероз и сосудистые органные поражения.
Материалы и методы
Обследуемые группы
В исследование были включены 2 группы сравнения:
1) группа риска ССЗ из 101 ребенка и подростка в возрасте от 4-х до 17-ти лет из семей, имеющих наследственную отягощенность по сердечно-сосудистой патологии у родственников первой и второй степеней родства;
2) контрольная группа из 144 практически здоровых детей и подростков сопоставимого возраста, не имеющих наследственной отягощенности по ССЗ и связанных с ними осложнений.

Электрофорез
Это метод разделения смеси веществ (белков, нуклеиновых кислот) на ее составляющие по массе в электрическом поле на носителе. В качестве носителя используют гель из агарозы. Концентрация агарозы в геле варьируется в зависимости от массы разделяемых фрагментов. В состав геля также входят вода и реакционный буфер. Расплав агарозы хранится в термостате при 60 градусах. Гель получается при заливке этого расплава на подложку, где он, при комнатной температуре, застывает В застывшем геле делаются лунки. Такой гель помещается в кювету между электродами. Сама кювета тоже заполнена буфером. Проба смешивается с краской и наносится в лунки геля. Затем в электрическом поле происходит разгонка пробы. Каждая фракция пробы с одинаковой массой формирует полосу. Эта полоса светится красным в ультрафиолете, за счет красителя. Определение массы этой фракции проводится относительно маркера. Маркер при разгонке в электрическом поле дает полосы с известной массой.
Параметры электрофореза: 120В, 70мА, 90 минут
1 2 3 4 5 6 7 8 9
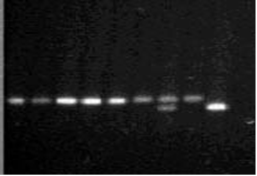
Рисунок 2
a4b полиморфизм гена eNOS, электрофорез в 2% агарозе.
, 2, 3, 4, 5, 6, 8 - норма 4b/4b (420 п.н.)
- гетерозигота 4а/4b (420, 393 п. н.)
- мутация 4а/4а (393 п. н.)
Статистический анализ
Статистическая обработка данных проводилась с помощью теста Хи-квадрат, точного критерия Фишера и метода относительного риска (ОР). Различия между сравниваемыми группами считались статистически значительными для доверительного интервала (ДИ) 95% (p=0,05). Проверка равновесия распределения генотипов в исследованных выборках, включая и случаи множественных сравнений внутри выборок, с законом Харди-Вайнберга проводилась с помощью теста Хи-квадрат.
Результаты и обсуждение
Распределения частот встречаемости аллелей и генотипов eNOS 4a/4b в изученных группах детей, представлены в таблице I. Частоты встречаемости аллелей и генотипов в исследуемых выборках не отличались значимо от теоретически ожидаемых в соответствии с законом Харди-Вайнберга.
Таблица I. Частоты встречаемости аллелей и генотипов eNOS 4a/4b в исследуемых группах детей
|
| мальчики
| девочки
| общее
|
|
| Группа риска
| Контрольная группа
| Группа риска
| Контрольная группа
| Группа риска
| Контрольная группа
|
| 4a/4b
| 12
| 17,14%
| 25
| 32,89%
| 10
| 32,26%
| 19
| 27,95%
| 22
| 44
|
| 4a/4a
| 1
| 1,43%
| 3
| 3,95%
| 2
| 6,45%
| 5
| 7,35%
| 3
| 8
|
| 4b/4b
| 57
| 81,43%
| 48
| 63,16%
| 19
| 61,29
| 44
| 64,70%
| 76
| 92
|
| 4a
| 14
| 10%
| 31
| 20,39%
| 14
| 22,58%
| 29
| 21,32%
| 28
| 60
|
| 4b
| 126
| 90%
| 121
| 79,61%
| 48
| 77,42%
| 107
| 78,68%
| 174
| 228
|
На первом этапе мы провели сравнительный анализ распределения частот генотипов eNOS 4a/4b между детьми группы риска и детьми контрольной группы. Мы не обнаружили статистически значимых отличий в распределении генотипов (Х2=3.697, р=0,157).
Затем мы провели сравнительный анализ распределения частот аллелей 4а и 4b в группе риска по сравнению с контрольной группой и также не нашли статистически значимых различий между нашими исследуемыми группами (Х2=3.458, р=0,063).
Затем мы провели сравнительный анализ распределений частот генотипов в исследуемых группах отдельно для мальчиков и отдельно для девочек. При этом мы объединили генотипы 4a/4a и 4a/4b в один класс, и сравнили частоты распределения генотипа 4b/4b и носителей 4а аллеля в виде 4a/4a или 4a/4b генотипов для корректного применения теста Хи-квадрат.
Сравнение распределений генотипов в группах мальчиков показало статистически значимые различия между мальчиками из группы риска и мальчиками из контрольной группы (Х2=5.152, р=0,023), которые выражались в том, что количество носителей 4а аллеля мальчиков контрольной группы было примерно в 2 раза больше, чем у мальчиков группы риска (ОР=1,98 для 95% ДИ 1,12 - 3,51). Также у мальчиков контрольной группы частота встречаемости аллеля 4а была примерно в 2 раза выше, чем у мальчиков группы риска (ОР=2,04 для 95% ДИ 1,13 - 3,67). Между девочками групп сравнения статистически значимых различий в частотах аллелей и генотипов обнаружено не было (Х2=0.010, р=0,918).
На последнем этапе мы провели статистический анализ распределения генотипов внутри группы риска в зависимости от типа наследственной отягощённости и встречаемости её в родословной семей обследуемых детей (таблицы II-IV).
Для начала мы сравнили распределения генотипов в группе риска в зависимости от количества у исследованных детей родственников с ожирением. Мы ранжировали детей групп риска на три класса. Класс (1-3) включал детей, имевших от 1 до 3х близких родственников с ожирением, класс (4-6)-соответственно 4-6 родственников с ожирением и класс (7-10) - от 7 до 10 родственников с ожирением (см. таблицу II).
сердечный полиморфизм ген наследственный дети
Таблица II. Частоты встречаемости генотипов eNOS 4a/4b в группе детей с риском ССЗ в зависимости от количества родственников с ожирением
| Количество родственников с ожирением
| Количество детей
| 4a/4a+4a/4b
| 4b/4b
|
| Класс (1-3)
| 42
| 8
| 34
|
| Класс (4-6)
| 37
| 13
| 24
|
| Класс (7-10)
| 8
| 2
| 6
|
Сравнительный анализ между классами был проведен с использованием точного критерия Фишера и не выявил статистически значимых различий в распределениях генотипов между классами (р=0,653). То есть распределение генотипов eNOS 4a/4b в группе детей с риском ССЗ не зависит от встречаемости в родословной родственников с ожирением.
Затем мы сравнили распределения генотипов в группе риска в зависимости от того, имели ли близкие родственники СД2 или не имели (см. таблицу III).
Таблица III. Частоты встречаемости генотипов eNOS 4a/4b в группе детей с риском ССЗ в зависимости от наличия родственников с СД2
| Наличие СД2
| Количество детей
| 4a/4a + 4a/4b
| 4b/4b
|
| Нет родственников с СД2
| 48
| 12
| 36
|
| Есть родственники с СД2
| 37
| 12
| 25
|
Сравнительный анализ также был проведен с использованием точного критерия Фишера и не выявил статистически значимых различий в распределениях генотипов eNOS 4a/4b в группе детей с риском ССЗ в зависимости от наличия у них родственников с СД2 (р=0,476).
Наконец, мы сравнили распределения генотипов в группе риска в зависимости от количества у исследованных детей близких родственников с ССЗ. Мы ранжировали детей группы риска на 4 класса в зависимости от количества родственников с ССЗ: класс (1-3),класс (4-6), класс (7-9) и класс (10-17) (см. таблицу IV).
Таблица IV. Частоты встречаемости генотипов eNOS 4a/4b в группе детей с риском ССЗ в зависимости от количества родственников с ССЗ
| Количество родственников с ССЗ
| Количество детей
| 4a/4a + 4a/4b
| 4b/4b
|
| Класс (1-3)
| 25
| 8
| 17
|
| Класс (4-6)
| 30
| 9
| 21
|
| Класс (7-10)
| 16
| 7
| 9
|
| Класс (10-17)
| 12
| 2
| 10
|
Затем мы методом точного критерия Фишера провели сравнение распределений генотипов между классами и также не обнаружили статистически значимых различий в зависимости от количества родственников с ССЗ. Группы были генетически однородны (р=0,223).
Таким образом, мы не обнаружили в исследованной выборке детей из группы риска различий в распределении генотипов и частот аллелей 4a/4b полиморфизма гена eNOS по сравнению с контрольной группой детей и в зависимости от типа отягощённой наследственности и её встречаемости в родословной детей. Присутствие аллеля 4а в генотипе детей из группы риска не было ассоциировано с присутствием конституциональных факторов риска развития ССЗ, а именно с отягощённой наследственностью по ожирению, СД2 и ССЗ. Более того, у мальчиков из группы риска количество носителей 4а аллеля было даже меньше, чем у мальчиков из контрольной группы. Наши результаты во многом согласуются с работами других авторов, которые также показывают отсутствие ассоциативных связей между 4a/4b полиморфизмом в гене eNOS и риском ССЗ. Однако существуют работы, которые обнаруживают ассоциативную связь полиморфизма гена eNOS с риском развития ССЗ, так как роль гена eNOS в системе эндотелиальной функции является ключевой. Поэтому в дальнейшем мы планируем изучить другие полиморфизмы в этом гене, в первую очередь С-786Т и Glu298Asp.
Выводы
) Сравнительный анализ распределения частот генотипов eNOS 4a/4b между детьми группы риска и детьми контрольной группы не выявил статистически значимых отличий в распределении генотипов
) Сравнительный анализ распределения частот аллелей 4а и 4b в группе риска по сравнению с контрольной группой также не выявил статистически значимых различий между нашими исследуемыми группами
) Сравнение распределений генотипов в группах мальчиков показало статистически значимые различия между мальчиками из группы риска и мальчиками из контрольной группы. Также у мальчиков контрольной группы частота встречаемости аллеля 4а была примерно в 2 раза выше, чем у мальчиков группы риска. Между девочками групп сравнения статистически значимых различий в частотах аллелей и генотипов обнаружено не было.
) Сравнение распределений генотипов eNOS 4a/4b в группе детей с риском ССЗ в зависимости от количества родственников с ожирением не выявило статистически значимых различий между нашими исследуемыми группами.
) Сравнение распределений генотипов eNOS 4a/4b в группе детей с риском ССЗ в зависимости от наличия родственников с СД2 не выявило статистически значимых различий между нашими исследуемыми группами.
) Сравнение распределений генотипов eNOS 4a/4b в группе детей с риском ССЗ в зависимости от количества родственников с ССЗ не выявило статистически значимых различий между нашими исследуемыми группами. Группы были генетически однородны.
СПИСОК ЛИТЕРАТУРЫ
1. М.В. Шестакова Дисфункция эндотелия - причина или следствие метаболического синдрома 2001.
2. Манухина Е. Б., Александров А. А. Оксид азота, дисфункция эндотелия и новые возможности патогенетической терапии сердечно-сосудистых заболеваний 2001.
. Минушкина Л., Затейщиков Д., Сидоренко Б. Генетические аспекты регуляции эндотелиальной функции при артериальной гипертонии Кардиология. 2000, №3, С. 68-76
. Lowenstein C.J., Glatt C.E., Bredt D.S., Snyder S.H. Cloned and expressed macrophage nitric oxide synthase contasts with the brain enzyme // Proc. Natl. Acad. Sci. USA. - 1992. - Vol. 89. - P. 6711-6715.
. Miyahara K., Kawamoto T., Sase K., et al. Cloning and structural characterization of the human endothelial nitric-oxide-synthase gene. Eur J Biochem. 1994 Aug 1;223(3):719-26
. Cai H., Wilcken D.E., Wang X.L. The Glu298-->Asp (894G-->T) mutation at exon 7 of the endothelial nitric oxide synthase gene and coronary artery disease. J Mol Med. 1999 Jun;77(6):511-4.
. Tomasz J. Guzik,1 Edward Black., et al. Relationship Between the G894T Polymorphism (Glu 298 Asp Variant) in Endothelial Nitric Oxide Synthase and Nitric Oxide-Mediated Endothelial Function in Human Atherosclerosis
. Nakayama M., Yasue H., Yoshimura M. et al. T-786®C mutation in the 5’-flanking region of the endothelial nitric oxide synthase gene is associated with coronary spasm // Circulation. - 1999. - Vol. 99. - P. 2864-2870.
9. Ichihara S., Yamada Y., Fujimura T., et al. Association of a polymorphism of the endothelial constitutive nitric oxide synthase gene with myocardial infarction in the Japanese population. Am J Cardiol. 1998 Jan 1;81(1):83-6.
10. Ignarro L.J. Biological actions and properties of endothelium-derived nitric oxide formed and released from artery and vein. Circ Res. 1989 Jul;65(1):1-21.
11. Poirier O., Mao C., Mallet C., et al. Polymorphisms of the endothelial nitric oxide synthase gene - no consistent association with myocardial infarction in the ECTIM study. Eur J Clin Invest. 1999 Apr;29(4):284-90.
12. Tsukada T., Yokoyama K., Arai T., et al. Evidence of association of the ecNOS gene polymorphism with plasma NO metabolite levels in humans. Biochem Biophys Res Commun. 1998 Apr 7;245(1):190-3.
. Wang H.U., Anderson D.J. Eph family transmembrane ligands can mediate repulsive guidance of trunk neural crest migration and motor axon outgrowth. Neuron. 1997;18:383-396.
. Britten M.B., Schachinger V., Dimmeler S., et al. ecNOS-polymorphism is associated with coronary endothelial disfunction. Eur Heart J 1999;20(144):907.
. Hou L., Osei-Hyiaman D., Yu H., et al. Association of a 27-bp repeat polymorphism in ecNOS gene with ischemic stroke in Chinese patients. Neurology. 2001 Feb 27;56(4):490-6.
. Чистяков Д.А., Воронько О.Е., Савостьянов К.В. и др. Полиморфные маркеры генов эндотелиальной NO-синтазы и сосудистого рецептора ангиотензиногена II и предрасположенность к ишемической болезни сердца. Генетика. 1999
. Котовская Ю.В., Кобалаева Ж.Д., Сергеева Т.В. и др. Полиморфизм генов ренин-ангиотензиновой системы и гена эндотелиальной NO-синтазы и макрососудистые осложнения при сахарном диабете типа 2. Артериальная гипертензия. 2002. Т.8, *3. С.86-90.
. Nakagami H., Ikeda U., Maeda Y., et al. Coronary artery disease and endothelial nitric oxide synthase and angiotensin-converting enzyme gene polymorphisms. J Thromb Thrombolysis. 1999 Oct;8(3):191-5.
. Granath B., Taylor R.R., van Bockxmeer F.M., Mamotte C.D. Lack of evidence for association between endothelial nitric oxide synthase gene polymorphisms and coronary artery disease in the Australian Caucasian population. J Cardiovasc Risk. 2001 Aug;8(4):235-41.
. Yahashi Y., Kario K., Shimada K., Matsuo M. The 27-bp repeat polymorphism in intron 4 of the endothelial cell nitric oxide synthase gene and ischemic stroke in a Japanese population. Blood Coagul Fibrinolysis. 1998 Jul;9(5):405-9.
. Zhao Qi, SU Shao-yong, CHEN Shu-feng., et al. Association study of the endothelial nitric oxide synthase gene polymorphisms with essential hypertension in northern Han Chinese 2006.
. Taniwaki H, Ishimura E, Matsumoto N, Emoto M, Inaba M, Nishizava Y Relations Between ACE Gene and ecNOSGene Polymorphisms and Resistive Index in Type 2 Diabetic Patients With Nephropathy //Diabets Care. 2001; 24: 1653-1660.
Список сокращений
Введение
Обзор литературысинтетаза и атерогенез






