СУПЕРСПИРАЛИЗАЦИЯ ДНК — структурная форма двуцепочечной молекулы ДНК, в которой она образует супервитки, пересекая при этом собственную ось. Суперспирализованная ДНК обладает большой энергией. В нормальных условиях в ДНК образуются только отрицательные супервитки, которые закручивают ДНК вокруг ее оси против часовой стрелки, т.е. в направлении, противоположном правосторонней двойной спирали. Имея форму суперспирали, ДНК в большей степени подвержена структурным изменениям. Гистоны и строение хроматина.Хроматин — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК.
Основную массу хроматина составляют белки гистоны. Гистоны являются компонентом нуклеосом, — надмолекулярных структур, участвующих в упаковке хромосом. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: H2A, H2B, H3 и H4. В одну нуклеосому входят по два белка каждого типа — всего восемь белков. Гистон H1, более крупный, чем другие гистоны, связывается с ДНК в месте её входа на нуклеосому. Нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно его называют конденсированным илигетерохроматином, он хорошо видим под микроскопом. ДНК, находящаяся в гетерохроматине не транскрибируется, обычно это состояние характерно для незначащих или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки.
Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием и фосфорилированием
Считается, что в ядре существуют так называемые функциональные домены хроматина (ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». Вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины.Структуру хроматина формирует элементарная фибрилла диаметром 10 нм. Для нее известны 4 уровня укладки в более сложные структуры. Важнейший этап в структурных исследованиях хроматина - открытие в 1973 осн. структурной единицы хроматина - нуклеосомы. Она состоит из универсальной "кор"-частицы, образованной ДНК (146 нуклеотидных пар), октамером из 4 гистонов (Н2А, Н2В, НЗ и Н4 - по две молекулы каждого) и линкерной ДНКпеременной длины (0-80 нуклеотидных пар), связанной с гистоном H1. Последовательность расположения гистоноввдоль молекулы ДНК имеет вид -Н3 — Н2А — Н2В — (Н4, Н3)2 — Н2В — Н2А — Н3. Согласно пространств. модели А. Клуга "кор"-частица выглядит как плоский диск диаметром 11 нм, толщиной 5,7 нм, с осью симметрии 2-го порядка, на внеш. пов-сть к-рого навита двойная спираль ДНК в В-форме, образующая 1,75 витка левой суперспирали. Для фибриллы диаметром 10 нм предложена модель "бусы на нитке" со специфич. по отношению к нуклеотидной последовательности ДНК расположением нуклеосом (т. наз. фазированием). Следующий уровень организации представлен толстой фибриллой диаметром 30 нм. Ее описывают две альтернативные модели: регулярная спираль - соленоид, на один виток к-рой приходится от 3 до 7-8 нуклеосом и менее признанная глобулярная, где каждые 6-12 нуклеосом образуют глобулу. Важную роль в наднуклеосомной организации хроматина играет гистон H1. Детали устройства т. наз. петельной или доменной структуры хроматина и собственно хромосомы в метафазе (одна из стадий деления клетки) неизвестны. Интересна гипотеза о соответствии одного домена одному или, в крайнем случае, неск. генам. Гистоны — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНКв ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5, H2A, H2B, H3, H4. Гистоны H2A, H2B, H3, H4, называемых кóровымигистонами (от англ. core — сердцевина), формируютнуклеосому, представляющую собой белковую глобулу, вокруг которой накручена нить ДНК. Гистон H1/H5, называемый линкерным гистоном, связывается с внешней стороной нуклеосомы, фиксируя на ней нить ДНК. В хроматине гистоны составляют 25-40 % сухого веса. Благодаря высокому содержанию лизина иаргинина гистоны проявляют сильно оснóвные свойства. Гистоны непосредственно контактируют с ДНК и способны нейтрализовать отрицательный заряд фосфатных групп ДНК за счёт положительных зарядов аминокислотных остатков. Последовательность аминокислот в этих белках является консервативной и практически не различается в организмах различных таксонов. Гистоны присутствуют в ядрах эукариотических клеток; у бактерий гистонов нет, но они выявлены уархей группы Euryarchaea Физико-химические свойства ДНК: денатурация, ренатурация, вызкость, поглощение в УФ, реакционноспособность.Денатурация – это процесс нарушения высших уровней организации белковой молекулы (вторичного, третичного, четвертичного) под действием различных факторов.
Ренатурация — это полное или частичное восстановление денатурированными биополимерами своих естественных свойств, в т. ч. биологической активности.Для оценки перехода ДНК от нативного состояния к денатурированному используют ряд методов. Чаще всего измеряют поглощение в УФ-области. Интенсивность поглощения света пуриновыми и пиримидиновыми основаниями в расчете на одно основание зависит от того, присутствует ли оно в свободном состоянии или входит в состав полинуклеотидов. При образовании двухспиральной структуры ДНК поглощение света каждым основанием уменьшается (явление гипохромизма). Разрушение двухцепочечной структуры при денатурации уменьшает «экранированность» оснований, интенсивность поглощения УФ-света возрастает. Если относительное поглощение при 260 нм отложить по оси ординат, а температуру – по оси абсцисс, то получается S-образная кривая (профиль плавления).
Она показывает, что при нагревании ДНК ведет себя подобно кристаллам: двухцепочечная молекула «расплетается» на составляющие ее цепи в пределах небольшого температурного интервала. Поэтому денатурацию ДНК нередко называют плавлением, а температуру, при которой ДНК денатурирована на 50%, – температурой плавления – Tпл. S-образные профили плавления показывают, что денатурация ДНК – кооперативный процесс, т.е. каждое предшествующее изменение повышает вероятность последующего. Например, в результате внешнего воздействия разрывается несколько водородных связей между азотистыми основаниями, это приводит к нарушению стэкинг-взаимодействий, что облегчает разрыв следующих водородных связей, и т.д.; этот процесс можно сравнить с расстегиванием молнии
Основные виды РНК, их функции и локализация в клетке. Особенности строения видов РНК. Физико-химические свойства РНК. Молекулы РНК - это линейные полинуклеотидные цепи из нуклеотидов, соединенных между собой 5’, 3‘- фосфодиэфирными связями. Цепи полярны, то есть у них есть 5’- и 3‘-концы. Клетки содержат три основных типа РНК: рибосомную — рРНК, транспортную — тРНК и матричную (информационную) — мРНК. Каждая из этих РНК выполняет специфическую роль в сложном процессе биосинтеза белка, при котором последовательность аминокислот однозначно определяется нуклеотидной последовательностью ДНК.В эукариотических клетках существуют также малые ядерные РНК (мяРНК), являющиеся участниками процессинга РНК, и гетерогенные ядерные РНК (гяРНК), представляющие собой предшественников мРНК. Кроме того, обнаружена так называемая антисмысловая РНК, участвующая в регуляции процесса репликации ДНК.В процессе транскрипции нуклеотидная последовательность локуса (место в хромосоме, где находится ген) в ДНК копируется в молекулу РНК. Транскрибируются три вида генов. Транскрипты генов рРНК используются в синтезе рибосом, нуклеотидная последовательность мРНК переписывается в последовательность аминокислот при синтезе полипептида на рибосоме, а транскрипты генов тРНК связываются с аминокислотами, которые затем переносятся в рибосомный синтезирующий центр в последовательности, зашифрованной в мРНК; этот процесс называется трансляцией.
Рибосомная РНК. Она входит в состав клеточных органелл —рибосом. Биохимическая функция рРНК пока до конца не изучена. Предполагается, что она выполняет роль молекулярного каркаса, на котором крепятся участники процесса трансляции; рРНК имеет большую молекулярную массу (до 2 10  ), характеризуется метаболической стабильностью. На ее долю приходится до 85—90 % всех клеточных РНК. Степень спирализованности молекул рРНК находится в пределах 70—80 %.Предполагается, что в белоксинтезирующей системе клеткифункция рРНК не исчерпывается ролью структурного компонента. У прокариотов обнаружено, что в рРНК имеются небольшие участки, комплементарные участкам мРНК. Спаривание этих участков, видимо, способствует первоначальному связыванию мРНК с рибосомой. Не исключено, что некоторые участки рРНК играют определенную роль в формировании пептидтрансферазного центра рибосомы, ответственного за образование пептидных связей при синтезе белка.Транспортные РНК. Это низкомолекулярные нуклеиновые кислоты; молекулярная масса колеблется в пределах 23 000—30 000, каждой из 20 белковых аминокислот соответствует, по крайней мере, одна тРНК. Однако некоторым аминокислотам специфичны от 2 до 6 тРНК; предполагается их общее количество около 60. Они составляют примерно 15 % общего количества клеточных РНК. Многие тРНК получены в гомогенном состоянии, некоторые — в кристаллическом виде.
), характеризуется метаболической стабильностью. На ее долю приходится до 85—90 % всех клеточных РНК. Степень спирализованности молекул рРНК находится в пределах 70—80 %.Предполагается, что в белоксинтезирующей системе клеткифункция рРНК не исчерпывается ролью структурного компонента. У прокариотов обнаружено, что в рРНК имеются небольшие участки, комплементарные участкам мРНК. Спаривание этих участков, видимо, способствует первоначальному связыванию мРНК с рибосомой. Не исключено, что некоторые участки рРНК играют определенную роль в формировании пептидтрансферазного центра рибосомы, ответственного за образование пептидных связей при синтезе белка.Транспортные РНК. Это низкомолекулярные нуклеиновые кислоты; молекулярная масса колеблется в пределах 23 000—30 000, каждой из 20 белковых аминокислот соответствует, по крайней мере, одна тРНК. Однако некоторым аминокислотам специфичны от 2 до 6 тРНК; предполагается их общее количество около 60. Они составляют примерно 15 % общего количества клеточных РНК. Многие тРНК получены в гомогенном состоянии, некоторые — в кристаллическом виде.
Структура и физико-химические свойства РНК. Рибонуклеиновые кислоты (РНК) — однонитевые молекулы, поэтому в отличие от ДНК их вторичная и третичная структуры нерегулярны. Нуклеотидная цепь РНК обладает гибкой структурой, ее длина в зависимости от вида РНК может варьировать в очень широких пределах — от нескольких десятков до десятков тысяч нуклеотидных остатков; молекулярная масса РНК находится в пределах 10 —10 .
Последовательность нуклеотидных звеньев, соединенных фосфодиэфирной связью в неразветвленную полинуклеотидную цепь, представляет собой первичную структуру РНК. Вторичная и третичная структуры РНК, определяемые как пространственная конформация полинуклеотидной цепи, формируются в основном за счет водородных связей и гидрофобных взаимодействий между азотистыми основаниями. Если для молекулы нативной ДНК характерна устойчивая спираль, то структура РНК более многообразна и лабильна. Рентгеноструктурный анализ показал, что отдельные участки полинуклеотидной цепи РНК, перегибаясь, навиваются сами на себя с образованием внутриспиральных структур. Стабилизация структур достигается за счет комплементарных спариваний азотистых оснований антипараллельных участков цепи; специфическими парами здесь являются А - U, G - С и, реже, G – U.Образование спиральных структур сопровождается гипохромным эффектом — уменьшением оптической плотности образцов РНК при 260 нм. Разрушение этих структур происходит при понижении ионной силы раствора РНК или при его нагревании до 60—70 °С; оно также называется плавлением и объясняется структурным переходом спираль — хаотический клубок, что сопровождается увеличением оптической плотности раствора нуклеиновой кислоты.Хотя РНК относится к однониточным полинуклеотидам, вместе с тем в ее цепях имеются участки различной длины, состоящие из комплементарных друг другу нуклеотидных последовательностей, включающих от десятков до тысяч нуклеотидных остатков, расположенных на небольшом удалении друг от друга. Благодаря этому в молекуле РНК возникают как короткие, так и протяженные биспиральные участки, принадлежащие одной цепи; эти участки носят название шпилек. Модель вторичной структуры РНК со шпилькообразными элементами была создана в конце 50-х — начале 60-х гг. XX в. в лабораториях А. С. Спирина (Россия) и П. Доти (США). Особенности строения РНК Нуклеотиды РНК состоят из сахара - рибозы, к которой в положении 1 'присоединена одна из основ: аденин, гуанин, цитозин или урацил. Фосфатная группа объединяет рибозы в цепочку, образуя связи с 3 'атомом углерода одной рибозы и в 5' положении другого. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК - можно назвать полианионом. РНК транскрибируется как полимер четырех оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C)), но в «зрелой» РНК есть много модифицированных оснований и сахаров. Всего в РНК насчитывается около 100 различных видов модифицированных нуклеозидов, из которых: - 2'-О-метилрибоза наиболее частая модификация сахара; - Псевдоуридин - наиболее часто модифицированная основа, которая встречается чаще всего. В псевдоуридине (Ψ) связь между урацилом и рибозой не C - N, а C - C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК. Еще одной модифицированной основой, о которой стоит сказать является - гипоксантин, деаминованний гуанин, нуклеозид которого носит название инозин. Инозин играет важную роль в обеспечении вырожденности генетического кода. Роль многих других модификаций не до конца изучена, но в рибосомальной РНК много пост-транскрипционных модификаций находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующих в образовании пептидной связи. Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырех нуклеотидов, в которой есть пара оснований аденин - гуанин. Важная структурная особенность РНК, отличающая ее от ДНК - наличие гидроксильной группы в 2 'положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, которая наиболее часто наблюдается в ДНК. В А-форме глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка. Второе последствие наличия 2 'гидроксильной группы состоит в том, что конформационно пластичные, то есть, не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять. «Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуется посредством водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры - стебель-петли, петли и псевдоузлы. В силу большого количества возможных вариантов спаривания оснований, предсказания вторичной структуры РНК - гораздо более сложная задача, чем структуры белков, но в настоящее время есть эффективные программы. Локализация ДНК и РНК в клетке различна. ДНК локализована в ядре клетки, РНК – в ядре и цитоплазме. Только тРНК находится преимущественно к цитоплазме в растворенном виде. Как правило, нуклеиновые кислоты (ДНК и РНК) в живой клетке находятся в форме нуклеопротеинов. Основные структуры, содержащие нуклеопротеины, - это хроматин, где локализована ДНК, и рибосомы, где локализована РНК.В клетке встречаются также полирибосомы – образования, состоящие из вытянутой молекулы мРНК, окруженной глобулярными белками, на которую нанизаны рибосомы. Характер связей, стабилизирующих двуспиральную структуру ДНК.
Водородные связи:

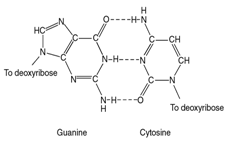
Стэкинг – взаимодействие:









