В основе возникновения электрических явлений в сердце лежит, как известно, проникновение ионов калия (К+), натрия (Na+), кальция (Са2+), хлора (С1~) и других через мембрану мышечной клетки. В электрохимическом отношении клеточная мембрана представляет собой оболочку, обладающую разной проницаемостью для различных ионов. Она как бы разделяет два раствора электролитов, существенно отличающихся по своему составу. Внутри клетки, находящейся в невозбужденном состоянии, концентрация К+ в 30 раз выше, чем во внеклеточной жидкости (рис. 1.1, а). Наоборот, во внеклеточной среде примерно в 20 раз выше концентрация Na+, в 13 раз выше концентрация С1 и в 25 раз выше концентрация Са2+ по сравнению с внутриклеточной средой. Такие высокие градиенты концентрации ионов по обе стороны мембраны поддерживаются благодаря функционированию в ней ионных насосов, с помощью которых ионы Na+, Ca2+ и С1- выводятся из клетки, а ионы К+ входят внутрь клетки. Этот процесс осуществляется против концентрационных градиентов этих ионов и требует затраты энергии.
В невозбужденной клетке мембрана более проницаема для К+ и С1-. Поэтому ионы К+ в силу концентрационного градиента стремятся выйти из клетки, перенося свой положительный заряд во внеклеточную среду. Ионы С1~, наоборот, входят внутрь клетки, увеличивая тем самым отрицательный заряд внутриклеточной жидкости. Это перемещение ионов и приводит к поляризации клеточной мембраны невозбужденной клетки: наружная ее поверхность становится положительной, а внутренняя — отрицательной (рис. 1.1,6). Возникающая таким образом на мембране разность потенциалов препятствует дальнейшему перемещению ионов (К+ — из клетки и С1- — в клетку), и наступает стабильное состояние поляризации мембраны клеток сократительного миокарда в период диастолы. Если мы теперь с помощью микроэлектродов измерим разность потенциалов между наружной и внутренней поверхностью клеточной мембраны, как это показано на рисунке 1.1, в, то зарегистрируем так называемый трансмембранный
| |
| |  |
| Глава 1. Биоэлектрические основы электрокардиографии
|
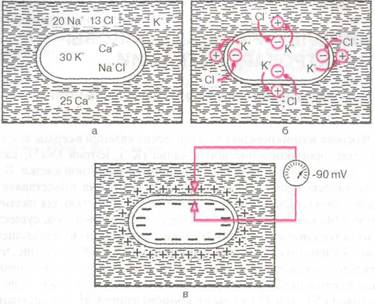
Рис. 1.1. Поляризация клеточной мембраны невозбужденной клетки: а — соотношение концентрации ионов Na+, K+, Cl- и Са2+ внутри клетки и во внеклеточной жидкости; б — перемещение ионов К+ и С1- вследствие концентрационного градиента; в — регистрация трансмембранного потенциала покоя
потенциал покоя (ТМПП), имеющий отрицательную величину, в норме составляющую около —90 mV
При возбуждении клетки резко изменяется проницаемость ее стенки по отношению к ионам различных типов. Это приводит к изменению ионных потоков через клеточную мембрану и, следовательно, к изменению величины самого ТМПП. Кривая изменения трансмембранного потенциала во время возбуждения получила название трансмембранного потенциала действия (ТМПД), Различают несколько фаз ТМПД миокар-диальной клетки (рис. 1.2).
Фаза 0. Во время этой начальной фазы возбуждения — фазы деполяризации — резко увеличивается проницаемость мембраны клетки для ионов Na+, которые быстро устремляются внутрь клетки (быстрый натриевый ток). При этом, естественно, меняется заряд мембраны: внутренняя поверхность мембраны становится положительной, а наружная — отрицательной. Величина ТМПД изменяется от —90 mV до +20 mV, т.е. происходит реверсия заряда — перезарядка мембраны. Продолжительность этой фазы не превышает 10 мс.
| 1.1. Мембранная теория возникновения биопотенциалов 13
|

Рис. 1.2. Трансмембранный потенциал действия (ТМПД). Объяснение в тексте. АРП и ОРП — абсолютный и относительный рефрактерные периоды
Фаза 1. Как только величина ТМПД достигнет примерно +20 mV, проницаемость мембраны для Na+ уменьшается, а для С1 увеличивается. Это приводит к возникновению небольшого тока отрицательных ионов С1- внутрь клетки, которые частично нейтрализуют избыток положительных ионов Na внутри клетки, что ведет к некоторому падению ТМПД примерно до 0 или ниже. Эта фаза носит название фазы начальной быстрой реполяризации.
Фаза 2. В течение этой фазы величина ТМПД поддерживается примерно на одном уровне, что приводит к формированию на кривой ТМПД своеобразного плато. Постоянный уровень величины ТМПД поддерживается при этом за счет медленного входящего тока Са2+ и Na+, направленного внутрь клетки, и тока К+ из клетки. Продолжительность этой фазы велика и составляет около 200 мс. В течение фазы 2 мышечная клетка остается в возбужденном состоянии, начало ее характеризуется деполяризацией, окончание — реполяризацией мембраны.
Фаза 3. К началу фазы 3 резко уменьшается проницаемость клеточной мембраны для Na+ и Са2+ и значительно возрастает проницаемость ее для К+. Поэтому вновь начинает преобладать перемещение ионов К+ наружу из клетки, что приводит к восстановлению прежней поляризации клеточной мембраны, имевшей место в состоянии покоя: наружная ее поверхность вновь оказывается заряженной положительно, а внутренняя поверхность - отрицательно.

 14 Глава 1. Биоэлектрические основы электрокардиографии
14 Глава 1. Биоэлектрические основы электрокардиографии
 ТМПД достигает величины ТМПП. Эта фаза носит название фазы конечной быстрой реполяризации.
ТМПД достигает величины ТМПП. Эта фаза носит название фазы конечной быстрой реполяризации.
Фаза 4. Во время этой фазы ТМПД, называемой фазой диастолы, происходит восстановление исходной концентрации К+, Na+, Ca2+, С1- соответственно внутри и вне клетки благодаря действию «Na+—К+-насоса». При этом уровень ТМПД мышечных клеток остается на уровне примерно —90 mV.
Клетки проводящей системы сердца и клетки синусового узла обладают способностью к спонтанному медленному увеличению ТМПП — уменьшению отрицательного заряда внутренней поверхности мембраны во время фазы 4. Этот процесс получил название спонтанной диастолической деполяризации и лежит в основе автоматической активности клеток синоатриального (синусового) узла и проводящей системы сердца, т.е. способности к «самопроизвольному» зарождению в них электрического импульса (подробнее см. ниже).
 Запомните!
Запомните!
Наружная поверхность клеточной мембраны заряжена:
1) положительно — в невозбужденной мышечной клетке, нахо
дящейся в состоянии покоя;
2) отрицательно — в клетке, находящейся в состоянии возбуж
дения в фазе 0 и 1 ТМПД (деполяризация и ранняя быстрая репо-
ляризация);
3) положительно — в клетке, восстанавливающей свой исход
ный потенциал (реполяризация клетки).
1.2. Основные функции сердца
Сердце обладает рядом функций, определяющих особенности его работы.
Функция автоматизма
Функция автоматизма заключается в способности сердца вырабатывать электрические импульсы при отсутствии внешних раздражений.
Функцией автоматизма обладают клетки синоатриального узла (СА-узла) и проводящей системы сердца: атриовентрикулярного соединения (АВ-соединения), проводящей системы предсердий и желудочков. Они получили название клеток водителей ритма - пейсмеке-ров (от англ. pacemaker — водитель). Сократительный миокард лишен функции автоматизма.
1.2. Основные функции сердца 15
Если в норме ТМПД сократительных мышечных клеток в течение всей диастолической фазы (фазы 4 ТМПД) стабильно поддерживается на одном и том же уровне, равном примерно -90 mV, для волокон водителей ритма (пейсмекеров) характерно медленное спонтанное уменьшение мембранного потенциала в диастолу, как это показано на рисунке
1.3. Этот процесс носит название медленной спонтанной диастолической
деполяризации и возникает в результате особых свойств мембраны пейс
мекеров — постепенного самопроизвольного увеличения в диастолу
проницаемости мембраны для ионов Na+, медленно входящих в клет
ку. В результате скопления в клетке все большего количества положи
тельных ионов отрицательный заряд внутренней поверхности клеточ
ной мембраны частично нейтрализуется и разность потенциалов между
наружной и внутренней поверхностью мембраны (ТМПП) постепенно
уменьшается. Как только ТМПП достигнет критического уровня (при
мерно -60 mV), проницаемость мембраны для ионов Na+ резко и быс
тро возрастает, что приводит к возникновению быстрой лавинообраз
ной деполяризации клетки (фаза 0 ТМПД) — ее возбуждению, которая
является импульсом к возбуждению других клеток миокарда.
Понятно, что чем выше скорость спонтанной диастолической деполяризации, тем чаще в клетках водителя ритма возникают электрические импульсы. В норме максимальной скоростью диастолической деполяризации и максимальной автоматической активностью обладают клетки СА-узла, который вырабатывает электрические импульсы с частотой около 60—80 в минуту. Это центр автоматизма первого порядка (рис. 1.4).

Рис. 1.3. Спонтанная диастолическая деполяризация волокон водителей
ритма — пейсмекеров. Объяснение в тексте.
а— ТМПД мышечных клеток; б — ТМПП клеток пейсмекеров

 16 Глава 1. Биоэлектрические основы электрокардиографии
16 Глава 1. Биоэлектрические основы электрокардиографии
 Функцией автоматизма обладают некоторые участки проводя-шей системы предсердий и АВ-соединение — зона перехода атрио-вентрикулярного узла (АВ-узла) в пучок Гиса' (см. рис. 1.4). Эти участки проводящей системы сердца, являющиеся центрами автоматизма второго порядка, могут продуцировать электрические импульсы с частотой 40—60 в минуту. Следует подчеркнуть, что сам АВ-узел, также входящий в состав АВ-соединения, не обладает функцией автоматизма.
Функцией автоматизма обладают некоторые участки проводя-шей системы предсердий и АВ-соединение — зона перехода атрио-вентрикулярного узла (АВ-узла) в пучок Гиса' (см. рис. 1.4). Эти участки проводящей системы сердца, являющиеся центрами автоматизма второго порядка, могут продуцировать электрические импульсы с частотой 40—60 в минуту. Следует подчеркнуть, что сам АВ-узел, также входящий в состав АВ-соединения, не обладает функцией автоматизма.
Наконец, центрами автоматизма третьего порядка, обладающими самой низкой способностью к автоматизму (25—45 импульсов в минуту), являются нижняя часть пучка Гиса, его ветви и волокна Пур-кинье2. Однако в норме возбуждение сердца происходит только в результате импульсов, возникающих в волокнах СА-узла, который является единственным нормальным водителем ритма. Дело в том, что в условиях сравнительно частой импульсации СА-узла подавляется автоматизм клеток АВ-соединения, пучка Гиса и волокон Пуркинье. Последние являются только потенциальными, или латентными, водителями ритма. При поражениях СА-узла функцию водителя ритма могут взять на себя нижележащие отделы проводящей системы сердца — центры автоматизма II и даже III порядка.
Запомните!
1. Все волокна проводящей системы сердца (кроме средней ча
сти АВ-узла) потенциально обладают функцией автоматизма.
2. В норме единственным водителем ритма является СА-узел,
который подавляет автоматическую активность остальных (экто
пических) водителей ритма сердца.
На функцию СА-узла и других водителей ритма большое влияние оказывает симпатическая и парасимпатическая нервная система: активация симпатической системы ведет к увеличению автоматизма клеток СА-узла и проводящей системы, а парасимпатической системы — к уменьшению их автоматизма.
1.2.2. Функция проводимости
Функция проводимости — это способность к проведению возбуждения, возникшего в каком-либо участке сердца, к другим отделам сердечной мышцы.
 ' По Международной анатомической номенклатуре — предсердно-желудочковый пучок. 2 По Международной гистологической номенклатуре — сердечный проводящий миоцит.
' По Международной анатомической номенклатуре — предсердно-желудочковый пучок. 2 По Международной гистологической номенклатуре — сердечный проводящий миоцит.

| 1.2. Основные функции сердца 17
|
Рис. 1.4. Проводящая система сердца. Объяснение в тексте
Функцией проводимости обладают как волокна специализированной проводящей системы сердца, так и сократительный миокард, однако в последнем случае скорость проведения электрического импульса значительно меньше.
Следует хорошо усвоить последовательность и особенности распространения возбуждения по различным отделам проводящей системы сердца. В норме волна возбуждения, генерированного в клетках СА-узла, распространяется по короткому проводящему пути на правое предсердие, по трем межузловым трактам — Бахмана, Венкебаха и Торе-ля — к АВ-узлу и по межпредсердному пучку Бахмана - на левое предсердие (см. рис. 1.4). Возбуждение распространяется по этим проводящим трактам в 2-3 раза быстрее, чем по миокарду предсердий. Общее направление движения волны возбуждения - сверху вниз и несколько влево от области СА-узла к верхней части АВ-узла. Вначале возбуждается правое предсердие, затем присоединяется левое, в конце возбуждается только левое предсердие (рис. 1.5). Скорость распространения возбуждения здесь невелика и составляет в среднем около 30—80 см • с"1. Время охвата волной возбуждения обоих предсердий не превышает 0,1 с.
 Запомните!
Запомните!
1. Направление распространения волны возбуждения по пред
сердиям — сверху вниз и немного влево.
2. Вначале возбуждается правoе, затем правое и левое предсер
дия, в конце — только левое предсердие.
3. Время охвата возбуждением предсердий не превышает в нор
ме 0,1 с.
| 18 Глава 1. Биоэлектрические основы электрокардиографии
|

Рис. 1.5. Распространение возбуждения по предсердиям: а — начальное возбуждение правого предсердия; б — возбуждение правого и левого предсердий; в — конечное возбуждение левого предсердия. Красным цветом показаны возбужденные (заштрихованные) и возбуждающиеся в настоящий момент (сплошные) участки. PI,, PII,PIII—моментные векторы деполяризации предсердий
В АВ-узле и особенно в пограничных участках между АВ-узлом и пучком Гиса происходит значительная задержка волны возбуждения, скорость проведения не более 2—5 см • с-'. Задержка возбуждения в АВ-узле способствует тому, что желудочки начинают возбуждаться только после окончания полноценного сокращения предсердий.
Малая скорость проведения электрического импульса в АВ-узле обусловливает и другую особенность его функционирования: АВ-узел может «пропустить» из предсердий в желудочки не более 180—220 импульсов в минуту. Поэтому при учащении сердечного ритма более 180—220 ударов в минуту некоторые импульсы из предсердий не достигают желудочков, наступает так называемая атриовентрикулярная блокада проведения. В этом отношении АВ-узел является одним из самых уязвимых отделов проводящей системы сердца.
 Запомните!
Запомните!
1. В АВ-узле происходит физиологическая задержка волны
возбуждения, определяющая нормальную временную последова
тельность возбуждения предсердий и желудочков.
2. При учащении сердечных импульсов, исходящих из СА-узла
или предсердий, более 180—220 в минуту даже у здорового челове
ка может наступить частичная атриовентрикулярная блокада про
ведения электрического импульса от предсердий к желудочкам.
 От АВ-узла волна возбуждения передается на хорошо развитую внут-рижелудочковую проводящую систему, состоящую из предсердно-желу-дочкового пучка (пучка Гиса), основных ветвей (ножек) пучка Гиса и волокон Пуркинье. В норме скорость проведения по пучку Гиса и его ветвям
От АВ-узла волна возбуждения передается на хорошо развитую внут-рижелудочковую проводящую систему, состоящую из предсердно-желу-дочкового пучка (пучка Гиса), основных ветвей (ножек) пучка Гиса и волокон Пуркинье. В норме скорость проведения по пучку Гиса и его ветвям

 1.2. Основные функции сердца 19
1.2. Основные функции сердца 19
 составляет 100—150 см • с-1, а по волокнам Пуркинье — 300—400 см. с-1. Большая скорость проведения электрического импульса по проводящей системе желудочков способствует почти одновременному охвату желудочков волной возбуждения и наиболее оптимальному и эффективному выбросу крови в аорту и легочную артерию. В норме общая продолжительность деполяризации желудочков колеблется от 0,08 до 0,10 с.
составляет 100—150 см • с-1, а по волокнам Пуркинье — 300—400 см. с-1. Большая скорость проведения электрического импульса по проводящей системе желудочков способствует почти одновременному охвату желудочков волной возбуждения и наиболее оптимальному и эффективному выбросу крови в аорту и легочную артерию. В норме общая продолжительность деполяризации желудочков колеблется от 0,08 до 0,10 с.
Для правильного понимания генеза различных зубцов ЭКГ необходимо хорошо знать нормальную последовательность охвата возбуждением (деполяризацией) миокарда желудочков. Поскольку волокна Пуркинье преимущественно располагаются в субэндокардиальных отделах желудочков, именно эти отделы возбуждаются первыми, и отсюда волна деполяризации распространяется к субэпикардиальным участкам сердечной мышцы (рис. 1.6). Процесс возбуждения желудочков начинается с деполяризации левой части межжелудочковой перегородки в средней ее трети (рис. 1.6, а). Фронт возбуждения при этом движется слева направо и быстро охватывает среднюю и нижнюю части межжелудочковой перегородки. Почти одновременно происходит возбуждение апикальной (верхушечной) области, передней, задней и боковой стенок правого, а затем и левого желудочка. Здесь возбуждение распространяется от эндокарда к эпикарду и волна деполяризации преимущественно ориентирована сверху вниз и вначале направо, а затем начинает отклоняться влево.
Через 0,04—0,05 с волна возбуждения уже охватывает большую часть миокарда левого желудочка, а именно его апикальную область, переднюю, заднюю и боковые стенки. Волна деполяризации при этом ориентирована сверху вниз и справа налево (рис. 1.6, б).
Последними в период 0,06—0,08 с возбуждаются базальные отделы левого и правого желудочков, а также межжелудочковой перегородки. При этом фронт волны возбуждения направлен вверх и слегка направо, как это показано на рисунке 1.6, в.
 Запомните!
Запомните!
1. В норме возбуждение распространяется по желудочкам за
0,08-0,10 с.
2. Волна деполяризации в стенке желудочка распространяется
от эндокарда к эпикарду.
3. Нормальная последовательность охвата возбуждением желу
дочков такова, что вначале деполяризуется межжелудочковая пе
регородка, затем большая часть правого и левого желудочков (вер
хушка, задняя и боковая стенки желудочков). Последними воз
буждаются базальные отделы левого и правого желудочков и меж
желудочковой перегородки.
| 20 Глава 1. Биоэлектрические основы электрокардиографии
|

Рис. 1.6. Распространение возбуждения по сократительному миокарду желудочков:
а — возбуждение (деполяризация) межжелудочковой перегородки (0,02с); б— деполяризация верхушек, передней, задней и боковой стенок желудочков (0,04—0,05с); в — деполяризация базальных отделов левого и правого желудочков и межжелудочковой перегородки (0,06—0,08 с). Цветовые обозначения те же, что и на рисунке 1.5
1.2.3. Функция возбудимости и рефрактерность волокон миокарда
Возбудимость — это способность сердца возбуждаться под влиянием импульсов. Функцией возбудимости обладают клетки как проводящей системы сердца, так и сократительного миокарда. Возбуждение сердечной мышцы сопровождается, как вы уже знаете (см. раздел 1.1), возникновением ТМПД и в конечном счете - электрического тока.
В разные фазы ТМПД возбудимость мышечного волокна при поступлении нового импульса различна. В начале ТМПД (фаза 0, 1, 2) клетки полностью невозбудимы, или рефрактерны, к дополнительному электрическому импульсу. Это так называемый абсолютный рефрактерный период миокардиального волокна, когда клетка вообще неспособна отвечать новой активацией на какой-либо дополнительный электрический стимул (см. рис. 1.2). В конце ТМПД (фаза 3) имеет место относительный рефрактерный период, во время которого нанесение очень сильного дополнительного стимула может привести к возникновению нового повторного возбуждения клетки, тогда как слабый импульс остается без ответа. Во время диастолы (фаза 4 ТМПД) полностью восстанавливается возбудимость миокардиального волокна, а его рефрактерность отсутствует.

 1.3. Формирование нормальной электрокардиограммы 21
1.3. Формирование нормальной электрокардиограммы 21
 1.2.4. Функция сократимости
1.2.4. Функция сократимости
Сократимость — это способность сердечной мышцы сокращаться в ответ на возбуждение. Этой функцией в основном обладает сократительный миокард. В результате последовательного сокращения различных отделов сердца и осуществляется основная - насосная - функция сердца.
1.3. Формирование нормальной электрокардиограммы
1.3.1. Формирование электрограммы одиночного мышечного волокна
Колебания величины ТМПД отражают динамику процессов де- и реполяризации в различных участках сердечной мышцы. Однако в клинической электрокардиографии электроды располагают на значительном удалении от миокардиальной клетки, и поэтому измерение ТМПД невозможно. Электрические потенциалы регистрируются обычно с поверхности возбудимой ткани или проводящей среды, окружающей сердце (эпикардиальной поверхности сердца, поверхности тела, конечностей, пищевода и т.д.).
Запомните!
Электрокардиограмма — запись колебаний разности потенциалов, возникающих на поверхности возбудимой ткани или окружающей сердце проводящей среды при распространении волны возбуждения по сердцу.
Разность потенциалов, создаваемая источником тока, характеризует напряжение, или электродвижущую силу (ЭДС), источника тока.
Вначале рассмотрим процесс формирования разности потенциалов на поверхности одиночного мышечного волокна и генез электрограммы (ЭГ) волокна (рис. 1.7). Как вам уже известно, в состоянии покоя вся наружная поверхность клеточной мембраны заряжена положительно. Между любыми двумя точками этой поверхности разность потенциалов отсутствует. На ЭГ одиночного мышечного волокна, зарегистрированной с помощью двух электродов, расположенных на поверхности клетки, записывается горизонтальная нулевая (изоэлектри-ческая) линия (рис. 1.7, а). При возбуждении миокардиального волокна (рис. 1.7, б) наружная поверхность деполяризованного участка заряжается отрицательно по отношению к поверхности участка, находящегося еще в состоянии покоя (поляризации); между ними появляет-
| 22 Глава 1. Биоэлектрические основы электрокардиографии
|

Рис. 1.7. Формирование разности потенциалов на поверхности одиночного мышечного волокна при его деполяризации и реполяризации и регистрация электрограммы (ЭГ) одиночного мышечного волокна. Объяснение в тексте. Красным цветом показаны возбужденные участки, стрелки обозначают направление движения волны деполяризации и реполяризации
ся разность потенциалов, которая и может быть зарегистрирована на ЭГ в виде положительного отклонения, направленного вверх от изолинии, — зубца R ЭКГ. Зубец R примерно соответствует фазе О ТМПД.
Когда все волокно окажется в состоянии возбуждения (рис. 1.7, в) и вся его поверхность будет заряжена отрицательно, разность потенциалов между электродами снова окажется равной нулю и на ЭГ будет записываться изолиния.



 1.3. Формирование нормальной электрокардиограммы 23
1.3. Формирование нормальной электрокардиограммы 23
 Запомните!
Запомните!
Быстрая деполяризация одиночного мышечного волокна на ЭГ, зарегистрированной с помощью поверхностных электродов, сопровождается быстрым положительным отклонением — зубцом R.
Далее в течение некоторого времени на ЭГ записывается горизонтальная, близкая к изоэлектрической, линия. Поскольку все участки миокардиального волокна находятся в фазе 2 ТМПД (фазе плато), поверхность волокна остается заряженной отрицательно, и разность потенциалов на поверхности мышечной клетки отсутствует или очень мала (см. рис. 1.7, в). Это сегмент RS— ТЭТ.
Запомните!
В течение времени, соответствующего полному охвату возбуждением волокна миокарда, на ЭГ регистрируется сегмент RS— T, в норме расположенный приблизительно на уровне изолинии.
 Процесс быстрой конечной реполяризации одиночного мышечного волокна (фаза 3 ТМПД) начинается в том же участке, что и волна деполяризации (рис. 1.7, г). При этом поверхность реполяризованно-го участка заряжается положительно, и между двумя электродами, расположенными на поверхности волокна, вновь возникает разность потенциалов, которая на ЭГ проявляется новым отклонением от изолинии — зубцом Т ЭГ. Поскольку к электроду, соединенному с «+» электрокардиографа, теперь обращена поверхность с отрицательным, а не с положительным зарядом, как при распространении волны деполяризации, на ЭГ будет регистрироваться не положительный, а отрицательный зубец Т. Кроме того, в связи с тем, что скорость распространения процесса реполяризации значительно меньше скорости перемещения фронта деполяризации, продолжительность зубца ГЭГ больше таковой зубца R, а амплитуда — меньше.
Процесс быстрой конечной реполяризации одиночного мышечного волокна (фаза 3 ТМПД) начинается в том же участке, что и волна деполяризации (рис. 1.7, г). При этом поверхность реполяризованно-го участка заряжается положительно, и между двумя электродами, расположенными на поверхности волокна, вновь возникает разность потенциалов, которая на ЭГ проявляется новым отклонением от изолинии — зубцом Т ЭГ. Поскольку к электроду, соединенному с «+» электрокардиографа, теперь обращена поверхность с отрицательным, а не с положительным зарядом, как при распространении волны деполяризации, на ЭГ будет регистрироваться не положительный, а отрицательный зубец Т. Кроме того, в связи с тем, что скорость распространения процесса реполяризации значительно меньше скорости перемещения фронта деполяризации, продолжительность зубца ГЭГ больше таковой зубца R, а амплитуда — меньше.
Запомните!
Процесс быстрой конечной реполяризации одиночного волокна на ЭГ регистрируется в виде отрицательного зубца Т.
Следует отметить, что на форму зубцов ЭГ влияет не только электрическая активность самого мышечного волокна, но и место расположения положительного и отрицательного электродов отведения, с помощью которого регистрируется ЭГ. Об этом и пойдет речь в следующем разделе.
1.3.2. Дипольные свойства волны деполяризации и реполяризации на поверхности одиночного мышечного волокна. Понятие о векторе
В клинической электрокардиографии электрические явления, возникающие на поверхности возбудимой среды (волокна, сердца), принято описывать с помощью так называемой дипольной концепции распространения возбуждения в миокарде. Это значительно упрощает трактовку всех электрокардиографических изменений, поэтому необходимо более подробно рассмотреть некоторые свойства сердечного диполя.
| Рис. 1.8. Направление вектора сердечного диполя при деполяризации (а) и реполяризации (б) одиночного мышечного волокна
|
Как видно на рисунке 1.8, процесс распространения волны деполяризации и волны реполяризации по одиночному мышечному волокну можно условно представить как перемещение двойного слоя зарядов, расположенных на границе возбужденного (-) и невозбужденного (+) участков волокна. Эти заряды, равные по величине и противоположные по знаку, находятся на бесконечно малом расстоянии друг от друга и обозначаются как элементарные сердечные диполи. Положительный полюс диполя (+) всегда обращен в сторону невозбужденного, а отрицательный полюс (—) - в сторону возбужденного участка миокардиального волокна. Диполь создает элементарную ЭДС.


 1.3. Формирование нормальной электрокардиограммы 25
1.3. Формирование нормальной электрокардиограммы 25
ЭДС диполя — векторная величина, которая характеризуется не только количественным значением потенциала, но и направлением — пространственной ориентацией от (—) к (+).
Запомните!
Условно принято считать, что вектор любого диполя направлен от его отрицательного полюса к положительному, как это показано на рисунке 1.8.
На рисунке также хорошо видно, что направление движения волны деполяризации по одиночному мышечному волокну всегда совпадает с направлением вектора диполя, а направление движения волны реполяризации противоположно ориентации вектора диполя.
Теперь, чтобы описать, как будет выглядеть форма ЭГ при любых направлениях движения волны де- и реполяризации, вам необходимо хорошо запомнить всего три общих правила.
Запомните!
Правило первое. Если в процессе распространения возбуждения вектор диполя направлен в сторону положительного электрода отведения, то на ЭГ мы получим отклонение вверх от изолинии — положительный зубец ЭГ (рис. 1.9, а).
Правило второе. Если вектор диполя направлен в сторону отрицательного электрода отведения, то на ЭГ мы зафиксируем отрицательное отклонение, вниз от изолинии, т.е. отрицательный зубец ЭГ (рис. 1.9,б).
Правило третье. Наконец, если вектор диполя расположен перпендикулярно к оси отведения, то на ЭГ записывается изолиния, т.е. отсутствуют положительные или отрицательные отклонения ЭГ (рис. 1.9, в).
 Эти простые правила позволят вам самостоятельно определить конфигурацию ЭГ при любом расположении активного положительного электрода и любом направлении движения волны де- и реполяризации, изображенных на рисунке 1.10. При решении этих заданий обязательно воспользуйтесь следующим алгоритмом:
Эти простые правила позволят вам самостоятельно определить конфигурацию ЭГ при любом расположении активного положительного электрода и любом направлении движения волны де- и реполяризации, изображенных на рисунке 1.10. При решении этих заданий обязательно воспользуйтесь следующим алгоритмом:
1) определите и отметьте на схеме полярность диполя во время де-
и реполяризации;
2) обозначьте стрелкой направление вектора диполя во время де- и
реполяризации;
3) схематично зарисуйте конфигурацию ЭГ во время де- и реполя
ризации.










