

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Топ:
Эволюция кровеносной системы позвоночных животных: Биологическая эволюция – необратимый процесс исторического развития живой природы...
Установка замедленного коксования: Чем выше температура и ниже давление, тем место разрыва углеродной цепи всё больше смещается к её концу и значительно возрастает...
Интересное:
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Распространение рака на другие отдаленные от желудка органы: Характерных симптомов рака желудка не существует. Выраженные симптомы появляются, когда опухоль...
Влияние предпринимательской среды на эффективное функционирование предприятия: Предпринимательская среда – это совокупность внешних и внутренних факторов, оказывающих влияние на функционирование фирмы...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
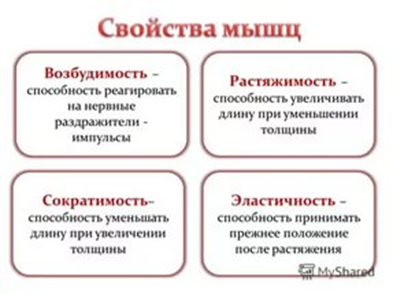
Скелетные мышцы. С помощью этих мышц тело удерживается в вертикальном положении, перемещается в пространстве, осуществляются дыхательные и глотательные движения, формируется мимика.
Гладкие мышцы. Мышцы непроизвольных движений, находятся главным образов в стенках полых органов, к примеру пищевода или мочевого пузыря. Короткие, веретеновидные мышцы, образующие пластины.
Сердечная группа мышц. Поперечнополосатая, исчерченная. Мышцы, находящиеся в сердце, отвечают за сердцебиение. 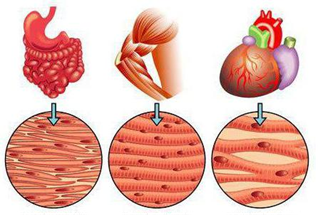
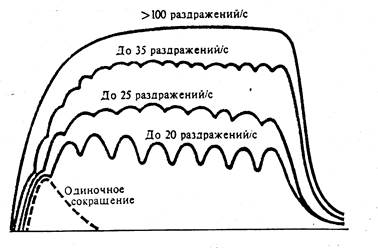
13. Одиночное сокращение скелетной мышцы.
Одиночное сокращение - это сокращение, которое возникает на одиночный стимул, достаточный для вызова возбуждения мышцы.
Одиночное сокращение мышечного волокна скелетной мышцы
Механический ответ отдельного мышечного волокна на одиночный потенциал действия называется одиночным сокращением (twitch). Основные характеристики одиночного изометрического сокращения показаны на рис. 30.20, а. Начало мышечного напряжения запаздывает на несколько миллисекунд по отношению к потенциалу действия. В течение этого латентного периода проходят все этапы электромеханического сопряжения.
Интервал от начала развития напряжения до момента его максимума - это время сокращения. Оно различно для разных типов волокон скелетных мышц. Время сокращения быстрых волокон не превышает 10 мс, тогда как для более медленных волокон оно не меньше 100 мс. Длительность сокращения определяется тем, как долго цитоплазматическая концентрация Са2+ остается повышенной, обеспечивая продолжение циклической активности поперечных мостиков. Время сокращения обусловлено активностью Са2+-АТФазы саркоплазматического ретикулума, которая в быстрых волокнах выше, чем в медленных.
|
|
Сравнение одиночных сокращений одного и того же мышечного волокна при разных режимах его деятельности показывает (рис. 30.20, б), что латентный период больше для изотонического сокращения, чем для изометрического, тогда как длительность механического процесса меньше в случае изотонического сокращения (т.е. при укорочении), чем изометрического (т.е. при генерировании силы).
Характеристики изотонического сокращения зависят также от веса поднимаемой нагрузки (рис. 30.21). А именно, при более тяжелой нагрузке:
- латентный период продолжительнее;
- скорость укорочения (величина укорочения мышцы в единицу времени), длительность сокращения и величина укорочения мышцы меньше.
Рассмотрим подробнее последовательность явлений во время изотонического одиночного сокращения. При возбуждении мышечного волокна поперечные мостики начинают развивать силу, однако укорочение не начнется, пока мышечное напряжение не превысит нагрузку на волокно. Таким образом, укорочению предшествует период изометрического сокращения, в течение которого возрастает напряжение. Чем тяжелее нагрузка, тем больше потребуется времени, чтобы оно сравнялось с величиной нагрузки и началось укорочение. Если нагрузку повышать, то в конце концов мышечное волокно не сможет ее поднять, скорость и степень укорочения будут равны нулю и сокращение станет чисто изометрическим.
 Изометрическое сокращение, зависимость между длиной мышечного волокна и его напряжением
Изометрическое сокращение, зависимость между длиной мышечного волокна и его напряжением Изометрические сокращения наблюдается при неизменной длине мышцы (например, при попытке поднять очень тяжёлый груз). В эксперименте этот вид сокращений можно получить, если жёстко закрепить мышцу с двух сторон и раздражать его.
|
|
Таким образом, условием изометрического сокращения является постоянная длина мышцы при максимальном тонусе.
7.Изотоническое сокращение, зависимость между скоростью сокращения мышцы и её нагрузкой
Изотоническое сокращение - это сокращение мышцы без нагрузки. Его можно получить в том случае, если мышцу закрепить с одной стороны, и раздражать её. В этом случае длина мышцы будет изменяться, а тонус окажется постоянным.
И при чисто изометрическом, и при чисто изотоническом сокращении работа совершаться не будет (нет перемещения груза). Поэтому в нормальных условиях скелетные мышцы сокращаются в смешанном (или ауксотоническом) режиме, где есть элементы и изометрического, и изотонического сокращения.
15. Закон силы и закон «все или ничего» в ответах возбудимых структур на раздражение.
Закон силы - зависимость силы ответной реакции ткани от силы раздражителя. Увеличение силы стимулов в определенном диапазоне сопровождается ростом величины ответной реакции. Чтобы возникло возбуждение, раздражитель должен быть достаточно сильным - пороговым или выше порогового. В изолированной мышце после появления видимых сокращений при достижении пороговой силы стимулов дальнейшее увеличение силы стимулов повышает амплитуду и силу мышечного сокращения. Действие гормона зависит от его концентрации в крови. Эффективность лечения антибиотиками зависит от введенной дозы препарата.
Закон все или ничего. Согласно этому закону, подпороговые раздражения не вызывают возбуждения ("ничего"), при пороговых и надпороговых стимулах возбуждение сразу приобретает максимальную величину ("все") и уже не увеличивается при дальнейшем усилении раздражения. По этому закону функционируют структурные единицы – мышечное волокно, нервное волокно.
16. ЭЭГ. Формирование суммарного электрического ответа нейронов.
Электроэнцефалограмма (ЭЭГ) - графическое изображение сложного колебательного электрического процесса, который регистрируется при помощи электроэнцефалографа при размещении его электродов на мозге или поверхности скальпа. С помощью ЭЭГ регистрируется суммарная электрическая активность головного мозга. В клинике обычно используется запись с помощью 10-12 электродов. Основными анализируемыми параметрами являются частота и амплитуда волновой активности.
|
|
Уровень возбуждения нейрона определяется суммой возбуждающих и тормозных воздействий, оказываемых на него в данный момент через синапсы. Если сумма возбуждающих воздействий больше суммы тормозных на величину, превышающую пороговый уровень, нейрон генерирует нервный импульс, распространяющийся затем по аксону. Описанным тормозным и возбуждающим процессам в нейроне и его отростках соответствуют определенной формы электрические потенциалы.
17. Роль синаптических и спайковых потенциалов в генерации ЭЭГ.
Синапти́ческие потенциа́лы, биоэлектрические потенциалы, возникающие в местах специализированных межклеточных контактов — синапсах — во время передачи возбуждения от одной клетки (пресинаптической) к другой (постсинаптической). Амплитуда и длительность синаптических потенциалов находятся под контролем внутриклеточных процессов и внешних воздействий. Снижение амплитуды возбуждающих синаптических потенциалов в определенных типах синапсов лежит в основе некоторых болезней человека (например, миастении, паркинсонизма).
Спайковый потенциал быстрое пикоподобное колебание электрического потенциала, сопровождающее возбуждение в нервных клетках. В физиологии термин «спайк» часто используют как синоним потенциала действия (ПД). В клинической электроэнцефалографии под спайковым потенциалом подразумевают характерные суммарные потенциалы длительностью до 10 мсек, внешне подобные ПД нерва, выявляемые в электрокортикограмме при некоторых патологических отклонениях, в частности при эпилепсии.
Спайковые потенциалы возникают в момент достижения мембраной нейрона некоторого критического уровня деполяризации, при котором наступает электрический пробой мембраны и начинается самоподдерживающийся процесс распространения возбуждения в нервном волокне.
|
|
При внутриклеточной регистрации спайк имеет вид высокоамплитудного, короткого, быстрого положительного пика.
Характерными особенностями спайков являются их высокая амплитуда (порядка 50-125 мВ), небольшая длительность (порядка 1-2 мс), приуроченность их возникновения к достаточно строго ограниченному электрическому состоянию мембраны нейрона (критический уровень деполяризации) и относительная стабильность амплитуды спайка для данногонейрона (закон все или ничего).
Частотный спектр ЭЭГ. Связь частотной структуры ЭЭГ с функциональным состоянием мозга.
Выделяют 5 главных ритмов, имеющих различные частотный диапазон, амплитуду и функциональное значение.
1. Дельта-ритм (0,5-4 Гц). Возникает при естественном и наркотическом сне, а так же наблюдается при регистрации ЭЭГ от участков коры, граничащих с областью, пораженных опухолью. У здорового взрослого человека практически отсутствует.
2. Тэта-ритм (5-7 Гц). Наиболее выражен в гиппокампе. Связан с поисковым поведением, усиливается при эмоциональном напряжении. Его иногда так и называют стресс-ритм. Сопровождается переживанием как положительных, так и отрицательных эмоций.
3. Альфа-ритм (8-13 Гц). Основной человеческий ритм, наблюдающийся в состоянии бодрствования, медитации и длительной монотонной деятельности. Наиболее выражен в затылочных областях головного мозга. Обычно эти колебания испытывают определенную модуляцию, так называемое «веретено альфа-ритма» – постепенное увеличение, а затем уменьшение амплитуды колебания альфа-ритма. Длительность веретен доля секунды-несколько секунд. «Десинхронизация» альфа-ритма – реакция пробуждения на раздражитель.
4. Мю-ритм — по частотно-амплитудным характеристикам сходен с альфа-ритмом, но преобладает в передних отделах коры больших полушарий.
5. Бета-ритм (15-35 Гц) – локализуется в предцентральной и фронтальной коре. Наблюдается при умственной активности у взрослых, причем значимое усиление высокочастотной активности наблюдается при умственной деятельности, включающей элементы новизны, в то время как стереотипные, повторяющиеся умственные операции сопровождаются ее снижением. Переход к состоянию напряжения сопровождается появлением бета-активности.
6. Гамма-ритм (выше 35 Гц). В предцентральной, теменной, височной зонах коры. Наблюдается при решении задач, требующих максимального сосредоточения.
18. Методики регистрации ЭЭГ.
Регистрация ЭЭГ производится при помощи электроэнцефалографа через специальные электроды (наиболее распространенные - мостиковые, чашечковые и игольчатые). В настоящее время чаще всего используется расположение электродов по международным системам «10—20 %» или «10—10 %». Каждый электрод подключен к усилителю. Для записи ЭЭГ может использоваться бумажная лента или сигнал может преобразовываться с помощью АЦП и записываться в файл на компьютере. ЭЭГ регистрирует разность потенциалов между двумя точками головы. Соответственно на каждый канал электроэнцефалографа подают напряжения, отведённые двумя электродами: одно на «вход 1», другое на «вход 2» канала усиления. Многоконтактный коммутатор отведений ЭЭГ позволяет коммутировать электроды по каждому каналу в нужной комбинации. Установив, например, на каком-либо канале соответствие затылочного электрода гнезду входной коробки «1», а височного - гнезду коробки «5», получают тем самым возможность регистрировать по этому каналу разность потенциалов между соответствующими электродами.
|
|
Метод вызванных потенциалов (ВП) представляет собой способ записи электрической активности групп нейронов спинного мозга, ствола мозга, зрительного бугра и больших полушарий после стимуляции той или иной афферентной системы зрительными, слуховыми или тактильными воздействиями. Амплитуда этих потенциалов, записываемых со скальпа с помощью обычных электроэнцефалографических электродов, колеблется от 0,5 и менее до 20 мкВ. В связи с их исключительно малой величиной они редко могут быть зарегистрированы на электроэнцефалографе с чернильными писчиками на фоне основной электрической активности мозга, которая обычно достигает по амплитуде 50 мкВ и более. Поэтому для извлечения из основной кривой ЭЭГ необходимо особое оборудование. Важно использовать особые усилители, с большой аккуратностью располагать электроды на поверхности скальпа, давать стимулы точно по времени и сводить к минимуму сопутствующие электрические артефакты. ВП расширяют возможности клинического неврологического исследования соответствующей афферентной системы, делают его более чувствительным и объективным, но не являются более специфичным методом этиологической диагностики.
19. Вызванные потенциалы мозга. Особенности регистрации ВП.
Вызванными потенциалами называются биоэлектрические сигналы, которые появляются с постоянными временными интервалами после определенных внешних воздействий. Для исследования ВП необходимо осуществить стимуляцию соответствующих рецепторов или нервных стволов, отвести электрические потенциалы от исследуемого отдела нервной системы, усилить их до необходимого уровня, подвергнуть определенной автоматической обработке в электронно-вычислительном устройстве и вывести в форме, удобной для анализа. Далее описаны основные технические данные устройств, необходимых для решения этих задач, и принципы их работы. Вызванный потенциал (ВП, EP от англ. evoked potential) - электрическая реакция мозга на раздражитель извне или на выполнение когнитивной задачи.
В практике используют визуальные, звуковые, электрические раздражители для регистрации зрительных, слуховых, соматосенсорных потенциалов соответственно. Регистрация осуществляется посредством электроэнцефалографических электродов, расположенных на поверхности головы.
Представления о генезе ВП. Ранние и поздние компоненты ВП.
ВП и последовательная модель переработки информации. Было установлено, что в конфигурации ВП можно выделять компоненты двух типов: ранние специфические (экзогенные) и поздние неспецифические (эндогенные) компоненты. Обработка физических параметров стимула (сенсорный анализ) связывается с деятельностью специфических сенсорных систем и находит свое отражение преимущественно в параметрах экзогенных компонентов ВП. В зрительной модальности эти компоненты регистрируются в интервале до 100 мс от начала ответа, в слуховой и соматосенсорной модальностях — еще раньше, в пределах 10-50 мс от начала стимуляции. Эндогенные компоненты ВП, возникающие позднее и длящиеся до окончания ответа, предположительно отражают этапы более сложной обработки стимула: формирование образа, сличение его с эталонами памяти, принятие перцептивного решения.
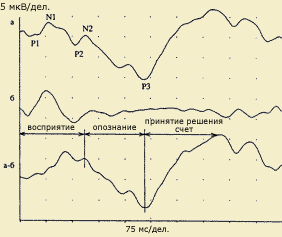
| Выделение когнитивной составляющей ответа: а - ответ в условиях опознания значимого стимула у здорового испытуемого (45 лет): P1-N1-P2 собственно ответ на стимул, N2 - 170 мс и P3 - 357 мс; б. Ответ на тот же стимул без опознания; а-б. Разность ответов в условиях распознавания и в стандартных условиях без распознавания. Когнитивный комплекс. Восприятие начинается на 76 мс, опознание и дифференцировка - компонент N2 - 170 мс, принятие решения и запоминание - компонент Р3 - 354 мс. (по В.В.Гнездицкому, 1997) |
Иллюстрацией такого подхода может служить концепция "информационного синтеза" А.М. Иваницкого (1986), согласно которой ранние компоненты ВП в интервале 0-100 мс связываются с оценкой физических параметров стимула, в среднелатентных компонентах в интервале 100-200 мс находят отражение процессы синтеза текущей сенсорной информации с содержанием памяти, а в поздних компонентах ВП отражается когнитивная оценка стимула как такового.
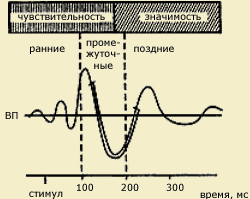
| Вызванные потенциалы (ВП) мозга, возникающие в ответ на световой или кожный раздражитель (по А.М.Иваницкому, 1999). Видно, что ранние волны соотносятся с чувствительностью, поздние - с мотивацией, значимостью стимула, а промежуточные - одновременно с двумя этими психофизическими показателями. Стрелкой на временной шкале обозначает момент предъявления стимула |
Эти представления хорошо согласуются с нейрокибернетическими построениями Д. Эделмена и В. Маунткасла (1981), согласно которым повторный возврат возбуждений является мозговой основой всех психических процессов. Так, например, момент возникновения образа совпадает по времени со встречей в коре двух информационных потоков, один из которых представляет собой внешний сигнал, а другой память. Предполагается, что эта память и есть возникающая в сознании человека частица "я", по отношению к которой сигнал воспринимается как нечто внешнее (см. Хрестомат. 5.3).
Изложенные выше представления объяснимы с помощью последовательной модели переработки информации, согласно которой каждая стадия перцептивного процесса наступает не ранее, чем реализуется предыдущая.
Сам вызванный потенциал - это сигнал прихода в кору определенной информации. Поэтому его анализ с информационной точки зрения не "фантазия", а естественная необходимость. Такой анализ может быть проведен на основании данных о генезе вызванного ответа и его отдельных волн.
20. Зрительные, слуховые, соматосенсорные ВП.
Зрительные вызванные потенциалы – биологические потенциалы, которые возникают в коре головного мозга в ответ на световое воздействие на сетчатку. В качестве стимулятора используют либо вспышки света, либо реверсивные шахматные патерны с экрана монитора.
В клинической практике наиболее востребованы коротколатентные акустические вызванные потенциалы, которые регистрируют потенциалы слухового нерва и акустических структур ствола головного мозга.
В качестве стимула могут использоваться тоны различной частоты или короткие щелчки с широким частотным спектром. Стандартно стимулы подаются через наушники.
Определение. ССВП — это электрический ответ центральной нервной системы на периферические сенсорные стимулы.
Выполнение ССВП позволяет выявить нарушения сенсорных путей.
|
|
|

Адаптации растений и животных к жизни в горах: Большое значение для жизни организмов в горах имеют степень расчленения, крутизна и экспозиционные различия склонов...

Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...

Двойное оплодотворение у цветковых растений: Оплодотворение - это процесс слияния мужской и женской половых клеток с образованием зиготы...

Таксономические единицы (категории) растений: Каждая система классификации состоит из определённых соподчиненных друг другу...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!